Hello everyone!! My name is Nadia Edelsztein and I am a PhD student from Argentina working in the Reproductive and Development field!
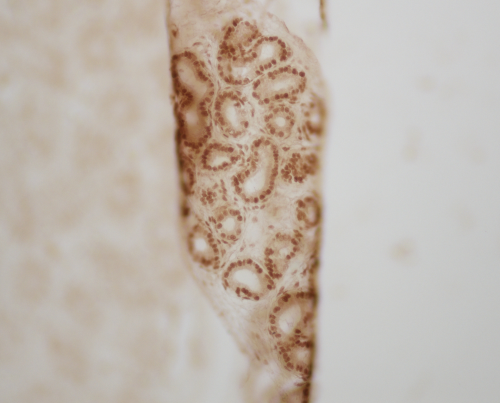
I am addressing you all to ask for a big favour. I entered a photo contest a couple of weeks ago, held by the institute where I used to learn German. The idea was to show pictures that, somehow, made you see things differently or from a different perspective. I chose a photo of one of the control immunohistochemistry assays I have done during my PhD project. It is an epididymis section from a 9dpp mouse that got all curled up during mounting but still looks beautiful -to me, at least. I have entitled it “Neben” (,,Nebenhoden” is epididymis in German). Even though I do not compete for the big prizes, I am in the run for “people’s choice award” and it is the only picture related to Science. You can only vote once and through Facebook.
So…I would like to ask you all if you could vote for my photo and, if you wish to do so, share the link with friends, acquaintances, etc! I am posting my pic at the end of this message for you to see which one is it.
In order to vote, you will have to click the following link and a website will open (it is in Spanish, so I apologise for that). There, you have to click on my photo (if you want to do so…please want to do so!!) and the little circle next to it turns black. Then go to the bottom of the page and click on the green button that says “Enviar” (which means “send” in Spanish). After that, there will be something displayed on a new page (all in Spanish) saying something very similar to “Thank you for participating! The winners will be announced July 5th, etc”.
3-D Retina Organoid Challenge to spur breakthroughs in treating blinding diseases
The National Eye Institute (NEI), part of the National Institutes of Health, has opened the first stage of a federal prize competition designed to generate miniature, lab-grown human retinas. The retina is the light- sensitive tissue in the back of the eye. Over the next three years pending availability of funds, NEI plans to offer more than $1 million in prize money to spur development of human retina organoids.
“None of the model systems currently available to researchers match the complex architecture and functionality of the human retina,” said NEI Director Paul A. Sieving, M.D., Ph.D. “We are looking for new ideas to create standardized, reproducible 3-D retina organoids that can speed the discovery of treatments for diseases such as age-related macular degeneration and diabetic eye disease, both leading causes of blindness.”
Currently, more than 4.2 million people over age 40 in the U.S. are visually impaired or blind, and that number is expected to double by the year 2050.1 Major visual disorders among Americans have an estimated annual economic burden of more than $35.4 billion.2
Research models are more valuable the more closely they mimic human tissue. Researchers hope to use retina organoids to study how retinal cells interact under healthy and diseased conditions, and to test potential therapies.
Stage I: Ideas
The ideation stage of the 3-D Retina Organoid Challenge aims to generate innovative ideas that can later be turned into concrete concepts. Running until August 1, 2017, the total prize purse for the ideation stage is $100,000.
“We’re looking for creative insights and application of new technology to unleash the full potential of retinal organoids. Our goal is for researchers to be able to generate or obtain retinal organoids easily so that they can be widely used for understanding diseases and testing drugs,” explained Jessica Mazerik, Ph.D., NEI challenge coordinator. “To do this, we are encouraging entries from diverse teams of participants.” They may come from vision research, developmental and stem cell biology, tissue engineering, materials science, 3-D bioprinting, and other fields.
Stage I also has a special solver category exclusively for trainees, which includes graduate and undergraduate students, postdoctoral fellows, and medical students. NEI has launched a discussion forum as a teambuilding space.
So far, nine sponsors have joined the challenge to support solvers through grants, access to expertise and discounted reagents, and in-kind testing.
Proposed Stage II: Development
The development stage of the challenge will require demonstration of a functional retina organoid prototype. This stage is planned to launch in fall 2017 and expected to offer $1 million in prize money.
Full details of the 3-D Retina Organoid Challenge prize competition are available at https://nei.nih.gov/3DROC.
References:
1. Varma R, Vajaranant TS, Burkemper B, Wu S, Torres M, Hsu C, Choudhury F, McKean-Cowdin R. Visual Impairment and Blindness in Adults in the United StatesDemographic and Geographic Variations From 2015 to 2050. JAMA Ophthalmol.134(7):802-809. doi:10.1001/jamaophthalmol.2016.1284 (2016).
2. Rein, D. B. et al. The economic burden of major adult visual disorders in the United States. Arch Ophthalmol. 124, 1754-1760. (2006).
You can also read a letter from the Director of the National Eye Institute, Paul Sieving, to Nature here:
NEI leads the federal government’s research on the visual system and eye diseases. NEI supports basic and clinical science programs to develop sight-saving treatments and address special needs of people with vision loss. For more information, visit https://www.nei.nih.gov.
About the National Institutes of Health (NIH): NIH, the nation’s medical research agency, includes 27 Institutes and Centers and is a component of the U.S. Department of Health and Human Services. NIH is the primary federal agency conducting and supporting basic, clinical, and translational medical research, and is investigating the causes, treatments, and cures for both common and rare diseases. For more information about NIH and its programs, visit https://www.nih.gov/(link is external).
Here are the highlights from the current issue of Development:
Btbd7 branches out across multiple organs
Dynamic changes in epithelial cell-cell adhesion and motility are crucial for branching morphogenesis – the process by which highly branched epithelial organs, such as the lung and kidney, grow and develop from a simple epithelial bud. Although many key regulatory factors involved in branching morphogenesis have been identified, it remains unclear how they coordinate to control epithelial cell dynamics.
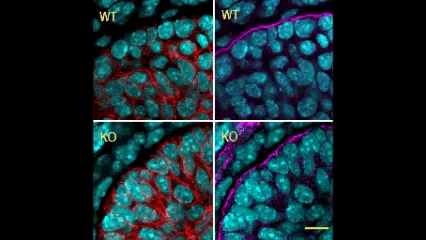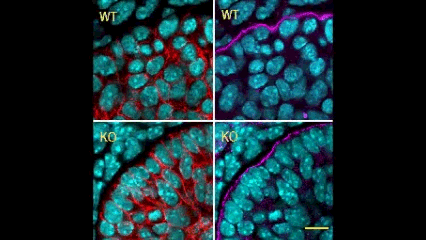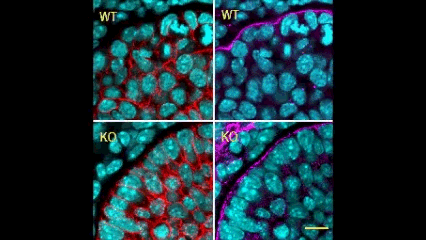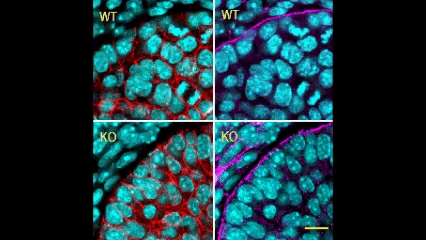
In this issue (p. 2200), the Yamada laboratory presents strong evidence for the essential role of Btbd7 in orchestrating epithelial remodelling during branching morphogenesis in the mouse lung, kidney and salivary glands in vivo. Specifically, the authors show that loss of Btbd7 in a knockout mouse model results in increased cadherin localisation to cell-cell junctions and reduced motility in the outer cells of epithelial buds. Conversely, Btbd7 overexpression in vitro leads to decreased cell-cell adhesion and increased cell motility. They further show that Btbd7 induces E-cadherin ubiquitination, internalisation and degradation in MDCK epithelial cells, providing a mechanism for how Btbd7 can control cell adhesion and migratory behaviour. Collectively, these data establish for the first time that the new regulatory molecule Btbd7 is required for successful in vivo branching morphogenesis of salivary gland, lung and kidney.
Female fertility: no bed of ROS(es)
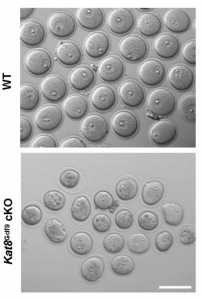
Oocyte development involves extensive transcriptional and epigenetic changes in order to eventually produce a mature egg competent for fertilisation. Key to this process is the avoidance of apoptosis, but how anti-apoptotic genes are regulated during oogenesis remains largely undefined. Now, on p. 2165, Qinghua Shi and colleagues show that genetic deletion of histone acetyltransferase KAT8 specifically in mouse oocytes at the primordial follicle stage causes the defective development of follicles from the secondary follicle stage, which subsequently leads to female infertility. The authors observe significantly increased reactive oxygen species (ROS) levels in the mutant oocytes, coincident with the downregulation of several antioxidant genes and show via chromatin immunoprecipitation assays that KAT8 regulates antioxidant gene expression by direct binding to promoter regions. Importantly, the authors are able to rescue the defects of folliculogenesis after Kat8 deletion in oocytes by antioxidant administration in mice. This study demonstrates for the first time that KAT8 represses ROS levels in oocytes by promoting the expression of antioxidant genes at the transcriptional level, and provides insight into the epigenetic regulation of female fertility.
Sox2+ pituitary stem cells on the MAP(K)
The MAPK/ERK pathway plays an important role in development and disease, with control over multiple cellular processes such as proliferation, differentiation and apoptosis, to name but a few. Despite its importance, little is known regarding its role in pituitary development and, specifically, in regulating the Sox2+ stem cell population therein. Now, on p. 2141, Scott Haston, from the Martinez-Barbera laboratory, and colleagues take a genetic approach to address the function of the MEK/ERK pathway during normal pituitary development, and relate this to human papillary craniopharyngioma (PCP), a form of benign but clinically relevant pituitary tumour. Using a MAPK gain-of-function mouse model, the authors demonstrate that constitutive activation of the MAPK/ERK pathway during pituitary development causes pituitary hyperplasia, abnormal morphogenesis and abnormal endocrine cell specification. This is due to sustained proliferation of the Sox2+ stem cell compartment, which results in an imbalance between proliferation and differentiation and, ultimately, an expansion of Sox2+ cells at the end of gestation. Looking at human PCP samples, the authors provide evidence that sustained proliferation of SOX2+ cells with reduced differentiation potential may contribute to the underlying pathogenesis of PCP. This study is an important step forward in understanding the role of the MAPK/ERK pathway in pituitary development, and sheds light on the possible pathogenesis of human PCP.
A new model for single-cell delamination
Epithelial cell delamination – the process by which individual epithelial cells detach from an epithelial layer – is a common phenomenon throughout development and can be observed across a wide range of species. Despite its prevalence, it remains unclear whether the mechanism that drives single-cell delamination varies according to the context and, if so, how.
In this issue (p. 2153), Yan Yan and colleagues investigate the mechanism of neuroblast delamination in the Drosophila embryo and find that it differs to that previously reported for epithelial homeostasis. Using an elegant combination of live imaging, genetics, pharmacology and mathematical modelling, the authors describe a new mechanism for cell delamination in which the neuroblasts undergo incremental apical constriction, which coincides with pulses of myosin accumulation at their medial apical cortex. By contrast, the accumulation of junctional myosin only has a weak correlation with apical constriction.
The authors demonstrate that the quantitative difference in the frequency and magnitude of myosin pulses critically determines whether a cell will effectively constrict, and further provide evidence for the possibility that this is regulated by the same signal that defines the neuroblast cell fate, namely, Notch. This study establishes a mechanism for cell delamination that is distinct from the previously reported mechanism, and opens up a new area of research into the possibility of Notch signalling upstream of dynamic cytoskeletal rearrangements.
MSPd signalling: new roles in muscle and reproduction
Amyotrophic lateral sclerosis (ALS), which is also known as Lou Gehrig’s disease and motor neurone disease (MND), is a debilitating disease that causes the death of both cortical and spinal motor neurons, resulting in a loss of control over voluntary muscle. There is currently no cure for ALS, and the origin of the disease is largely unknown. In previous studies, VAMP/synaptobrevin-associated proteins (VAPs) have been associated with ALS and spinal muscular atrophy (SMA): specifically, the N-terminal major sperm protein domain (MSPd), which serves as an extracellular signalling molecule and which is mutated in some forms of familial ALS. In this issue, Sung Min Han, Michael Miller and colleagues investigate the molecular framework and functional consequences of MSPd signalling in C. elegans, with implications for muscle and gonad development.
In the first study (p. 2175), the authors focus on the role of MSPd in muscle formation, based on the expression of VPR-1, the C. elegans homologue of human VAPB. The authors show that secreted MSPds promote the localisation of mitochondria within the body wall muscle during development, and that this is dependent on signalling events that involve the CLR-1 Lar-like phosphatase receptor. Using a targeted RNAi screen of known genes implicated in ALS and SMA, the authors identified survival of motor neuron 1 (SMN-1) as a crucial downstream mediator of MSPd and further showed that SMN-1 and ARX-2 are important for mitochondria localisation along the I-bands in body wall muscle.
In the second study (p. 2187), the authors turn their attention to the role of VPR-1 in gonad development, an area in which the role of VPR-1 remains largely unexplored. Taking a genetic approach, the authors show that vpr-1 null mutants are sterile upon hatching, a defect in gonadogenesis that can be rescued by the expression of MSPd from almost any tissue, except for the somatic gonad itself. The authors further demonstrate that, under normal conditions, gonad development depends on germline and neuronal expression of vpr-1 and specifically on cleaved MSPd, which circulates in the pseudocoelom and induces gonadogenesis non-cell-autonomously, much like a hormone.
Together, these two studies represent a major step forward in our understanding of the function of the vpr-1 gene in C. elegans, and also demonstrate the utility of this organism in elucidating disease mechanisms.
This Meeting Review of the 2017 RIKEN-CDB Symposium summarises recent progress in our understanding of human development, genetics and our evolutionary history.
This Review discusses the mechanisms that underpin the formation of skeletal muscle during development and its recapitulation from pluripotent stem cells in vitro.
This Review discusses the principal mechanisms involved in thyroid organogenesis, highlighting the factors involved in thyroid progenitor specification and the events occurring during thyroid gland morphogenesis.
Kamberov Laboratory, Department of Genetics in the Perelman School of Medicine at the University of Pennsylvania, USA.
Post-doctoral positions are available in the field of evolutionary and developmental
genetics.
Our lab’s research focuses on unraveling the developmental networks and genetic changes that underlie the divergence of humans from other species and for the diversity of present-day human populations. We are broadly interested in evolution of humans, but the core focus of the lab is on the skin and its appendages. We pursue these goals in order to not only understand the genetic origins of human specialization and variation but also seek to apply our findings to the betterment of human health. Available projects include: dissection of genetic pathways of skin appendage development and regeneration in mouse models; discovery and modeling of human adaptive variants using transgenic mice; high throughput screening for genetic elements controlling the development and uniqueness of human skin appendages.
A doctorate in biology or related field is required. Applicants with a strong background in genetics/genomics, developmental biology and molecular biology are encouraged to apply.
Interested candidates should provide: 1) your CV 2) A brief letter detailing your interest in the lab and relevant past research experience 3) The contact information for three references who can comment on your research. Application materials and any questions regarding the position should be sent to Yana Kamberov: yana2@mail.med.upenn.edu
Our image competition for a future Development cover is still live: voting closes tomorrow at 13.00 GMT! Click here to vote and find out more about the images
An NIH-funded postdoctoral position is available to investigate the signaling functions of retinoic acid (RA) during mouse embryo development. Our laboratory has reviewed recent advances in this field: Cunningham, T.J. and Duester, G. Mechanisms of retinoic acid signalling and its roles in organ and limb development. Nature Reviews Molecular Cell Biology 16: 110-123 (2015).
We are searching for a Postdoctoral Associate to explore the mechanisms of RA signaling and nuclear receptor coregulators during the early stages of organogenesis and limb formation using CRISPR/Cas9 gene editing, RNA-seq transcriptome analysis, and embryo chromatin immunoprecipitation.
Candidates should have a strong background in developmental biology or stem cell biology. Salary $47,476 with full benefits.
Interested applicants should email their CV and names of three references to:
Gregg Duester, Ph.D.
Professor
Development, Aging, and Regeneration Program
Sanford Burnham Prebys Medical Discovery Institute
Things are in full swing at MBL in the 2017 Embryology course, we are already one third of the way through! The first week we learned about echinoderms and C. elegans with a side of Tardigrade and this past we focused on zebrafish and Xenopus with axolotl. The atmosphere of the past two weeks has been full of excitement as we learn about new model systems and techniques. Not only did we discover new things scientifically, but we also learned more about each other as students and faculty.
A key element that appeared this week is our connection to each other and to the generation of scientists before us. It has been breath taking to learn about all the contributions that individuals and groups made to the field and to hear that so many of these discoveries were made either here at Woods Hole or by scientists who were a part of the course at some point during their career. It is thrilling to think that we are establishing meaningful connections, collaborations, and friendships that will be with us for the rest of our lives!
What enhances the connection is the unique diversity in the group, we all come from different places and have different backgrounds. The group is a mix of developmental, cell, and computational biologists as well as bioinformaticians, biophysicists and engineers. These differences in training and background paired with our general passion for science and inquiry have allowed us to look at problems from new and different angles. This is exciting, enriching and stimulates us as a group to propose and test new hypothesis.
The students in the Embryology course students come from all around the world!
(image made with travellerspoint formatted by Zuzka Vavrusova)
For up to date info on the course follow us on Twitter #embryo2017 or instagram #embryology2017
Insight into the organizational structure of a growing tissue is imperative for understanding its development and function. Structure can reveal the systematic steps undertaken towards making specific positional and cell fate choices/decisions. A well-defined structure helps to dissect the complexity underlying the networks that form as the tissue develops. It helps to elucidate the foundation on which the components can work together within the confines of the system. It helps one to read the lines of communication that are built into the system for its proper functioning. When an organized structure regulates the flow of information, then changes within that information are easier to monitor and adapt.
Historical views
The Enteric Nervous System (ENS) is one of the largest subdivisions of the peripheral nervous system. It earns its’ name as the “second brain” due to the millions of neurons it contains and the complex neural networks it displays1. Although Bayliss and Starling’s description of ‘the law of the intestine’ hallmarked the functional discovery of the ENS already more than a century ago,2 their discovery also unveiled some daunting scientific challenges, many of which are explained by inadequate insight into ENS structure and how this contributes to its function.
Simply put – It is a very challenging system for various different reasons. The ENS is challenged every day with the variety of food/drink we intake, which means the environment is under constant change. The ENS has a mind of its own! It autonomously controls functions of the gut that include peristalsis, secretion of enzymes and absorption of food. The ENS is layered in close apposition to contractile sheets of smooth muscle cell syncytia, thereby complicating several experimental approaches to a large extent. The gut lumen is host to a vast ocean of microbes with which the ENS interacts3 with other systems within this tissue, such as, the immune and epithelial systems to maintain a healthy gut. In addition, it also communicates with the brain to keep the body in good health.
The ENS originates from neural crest-derived progenitors that travel through different spatio-temporal environments as they expand in number, colonize the gut tissue and differentiate to generate a plethora of neuronal and glial cells. Unlike the central nervous system, neurons and glial cells of the ENS do not show any apparent order of arrangement. Cells coalesce in a salt and pepper manner to form ganglia and create an expansive network. However, reproducible patterns of secretory and motor function appear to be almost exclusively controlled by the ENS. Being indispensable for gut physiology, its lack of function is implicated in the pathogenesis of several gastrointestinal disorders, such as Hirschsprung disease and others that are of unknown aetiology.
Important questions
For a system that is tremendously interactive in an extensively expanding tissue (8 metres in humans), knowledge of how its structure develops is very important. This information can be useful in understanding diseases of the gut that may arise during development or later in adulthood and disrupt the network. Therefore, it was imperative for us to make sense of the ‘randomness’ that the ENS displays. We wished to gain an understanding of any fundamental rules that defined its apparent chaotic cellular topology and eventually its contribution to function.
Previous work in many labs including ours, have aimed to understand ENS structure and function by using global approaches and studying it at the population level. However, such approaches have not been very successful in gaining detailed insight into how the lineages of the ENS develop and contribute to its structure and function. As a lab that carries a reputation of developing tools and techniques that help to answer big over-arching questions in the field, we took the plunge. We decided to opt for single-cell approaches so as to address the contribution of an individual progenitor to the spatial development and function of the ENS at both the cellular and molecular level.
The work
Appropriate combination of genetic tools4 were generated in the lab and employed to track the static behaviour of individual ENS progenitors in a multi-colour mosaic manner over the developmental time line. Having optimized the various steps involved in the different experiments our results looked promising enough for me to spend hours and days to localize and image the beautiful clones we obtained in our study. The scanned clones were analysed for their composition, measured left, right and centre to gain an understanding about their spatio-temporal characteristics. We combined in vivo and ex-vivo approaches to capture the cellular properties of these cells. The days spent away from imaging were invested in isolating single cells for the transcriptomic study. This analysis was performed to understand the intrinsic properties of heterogeneous individual progenitors and how ENS lineages are generated. We also used mosaic mutagenesis to disrupt the system during development and understand its effect on ENS composition.
Making sense
With this arduous approach the mysterious topology of the ENS unfolded before our eyes. The close spatial relationship of the labelled families both at the 2D and 3D axes with a single colour meant ‘something’ to the system. This study helped us to unravel a set of rules that define the columnar organization of overlapping clonal lineages. This suggested to us that this structural organization could help the components of the ENS to work and face the challenges of the system together as a family. We also observed that as the system developed, subpopulations emerged in a defined manner. Neurons related to each other showed co-ordinate activity upon stimulation, highlighting their means of communication within the ENS of the small intestine. Molecular analysis of ENS progenitors revealed the manner in which neuronal and glial lineages arise. Further, our mutagenesis study revealed the specific role of an important receptor tyrosine kinase, RET in neuronal commitment. Together, our work suggests that lineage relationships are fundamental for the spatial organization and function of the ENS.
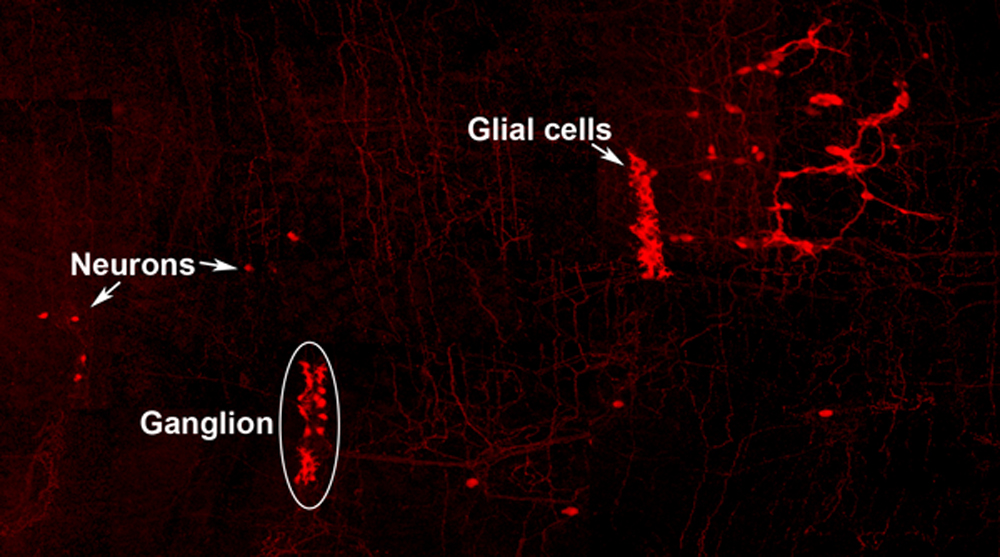
Neuro-glial clone: This image shows the progeny of a neuro-glial progenitor in the myenteric plexus of the adult mouse. Neurons extend processes forming a meshwork of connectivity and glial cells are observed within ganglia.
Future
Perhaps mistakes in the blueprint of the ENS are the cause of gastrointestinal diseases with unknown aetiology. We are yet to uncover the principles that underlie information processing, which will help us to assign the logic of ENS assembly and connectivity. Now that we have a better understanding of how the ENS of the small intestine assembles and underpins its function, we can start to probe this system at different stages of development.
This blog is contributed by Reena Lasrado and Werend Boesmans.
References
Bayliss WM, and Starling EH. The movements and innervation of the small intestine. The Journal of Physiology 24: 99-143, 1899.
Gershon MD. The Second Brain. New York, NY: HarperCollinsPublishers, 1998, p. xvi, 314 p.
We are seeking a highly motivated and ambitious candidate to join the research activities within our stem cell and developmental biology projects.
About us
DanStem comprises of two sections: The Novo Nordisk Foundation Section for Basic Stem Cell Biology that addresses basic research questions in stem cell and developmental biology (BasicStem). The Strategic Translational Stem Cell Research and Therapy (TransStem) Section that is focused on the translation of promising basic research results into new strategies and therapies to combat cancer and chronic diseases such as diabetes, Parkinson’s disease and liver failure. Find more information about the Center at http://danstem.ku.dk/
Job description
Using in vitro and in vivo models, we investigate how transcription factors and signaling pathways interact to define mesodermal cell fate in the early mouse embryo and thereby develop tools for maximizing protocols of stem cells differentiation. Specifically, we intend to dissect the mechanisms by which epiblast cells commit to different types of mesoderm progenitors. We intend of performing a high-throughput screen to identify transcription factors controlling the specification of different mesoderm populations.
A strong background in gene regulation, early mouse development and stem cell biology will be considered of particular relevance. Experience in bio-informatics would enhance the relevance of the candidate.
Qualifications
Candidates must hold a PhD degree in bioinformatics, genetics, stem cell biology, developmental biology, or bioengineering or in a relevant similar field.
An excellent publication record is required.
Experience with mouse/human ES cell culture will be an advantage.
An analytical aptitude and experience with high throughput screens is recommended.
Previous experience in bioinformatics, analysis of next-generation sequencing (NGS) data is considered of great advantage.
Knowledge in one of the scripting languages (Perl, Python, Shell) is advised.
Good English communication skills, both oral and written, are prerequisite for the successful candidate.
Terms of salary, work, and employment
The employment is for 2,5 years and is scheduled to start August 2017 or upon agreement with the chosen candidate. The place of work is the DanStem, University of Copenhagen, Blegdamsvej 3B, Copenhagen. Salary, pension and terms of employment are in accordance with the provisions of the collective agreement between the Danish Government and AC (the Danish Confederation of Professional Associations). In addition to the basic salary a monthly contribution to a pension fund is added (17.1% of the salary).
Curriculum vitae incl. education, experience, previous employments, language skills and other relevant skills
Copy of diplomas/degree certificate(s)
Three reference letters
How to apply
Your application must be submitted in English by clicking “Apply online”. Only online applications will be accepted.
The application will be assessed according to the Ministerial Order no. 284 of 25 April 2008 on the Appointment of Academic Staff at Universities.
The University of Copenhagen encourages all interested in the position to apply.
Application deadline: July 7th 2017
Only applications received in time and consisting of the above listed documents will be considered. Applications and/or any material received after deadline will not be taken into consideration
Founded in 1479, the University of Copenhagen is the oldest university in Denmark. It is among the largest universities in Scandinavia and is one of the highest ranking in Europe. The University´s eight faculties include Health Sciences, Humanities, Law, Life Sciences, Pharmaceutical Sciences, Science, Social Sciences and Theology. www.ku.dk
We have recently revised the model of Follicle Stem Cell (FSC) organization in the Drosophila ovary, showing that there is a much larger population of stem cells than formerly realized, that these FSCs exhibit population asymmetry, and that they give rise to Escort Cells as well as Follicle Cells [1]. Ovarian Germline Stem Cells have long been used as model stem cells, benefiting from easy recognition by virtue of their characteristic location at the anterior of the germarium, morphology, and functional markers that reflect the actions of a key BMP (bone morphogenetic protein) niche signal. FSCs, which are required for the development of germline cells into a mature oocyte, have been more difficult to investigate. No marker specific to these stem cells has been discovered, they are not in an easily recognized location and are not morphologically conspicuous among their neighbors.
In 1995 Margolis and Spradling reported the identification of FSCs (originally named Somatic Stem Cells), which they located midway through the germarium by BrdU labeling and lineage analysis [2]. An FSC is defined by lineage analysis as a cell that can produce Follicle Cells (FCs) but remains in the germarium while germline cysts pass through. FSC derivatives that associate with a germline cyst in the posterior half of the germarium all initially proliferate. A few quickly become specialized post-mitotic polar cells and stalk cells to allow egg chamber budding from the germarium, while the rest form an expanding epithelium around the germline cyst, cease divisions in mid-size egg chambers (stage 6) and adopt position-specific fates about a day later (stage 9). We refer here to all of these FSC derivatives including the earliest precursors (often termed prefollicle cells) associated with a germline cyst as Folicle Cells (FCs). Marked lineages that originate in an FC have a predictable, limited lifespan because all FC progeny move through the germarium and ovariole along with the associated cyst at an established rate. It takes the earliest FC about two days to exit the germarium and five days to exit the ovariole. If a marked cell remains in the germarium or the ovariole for longer than this time then we can deduce that the lineage must have originated in an FC precursor, namely an FSC. Although this definition has been used since 1995, our perception has changed from expecting most FSCs to reside in the germarium much longer than an FC, to realizing that many FSCs are in fact very short-lived before becoming FCs.
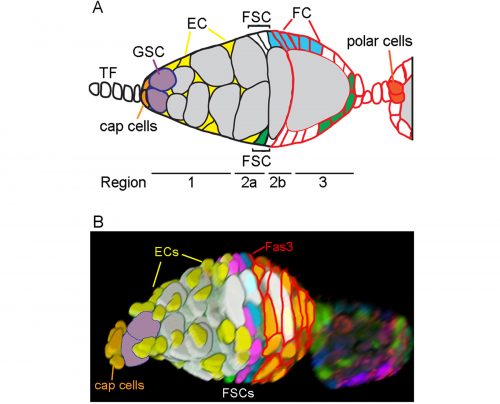
Location of FSCs in a germarium. A. Schematic showing terminal filament (TF) cells, Germline Stem Cells (GSC), GSC daughters developing into 16-cell cysts (grey), Escort Cells (EC, yellow), Follicle Stem Cells (FSC) and follicle cells (FC) from anterior (left) to the newest egg chamber. Two potential FSC lineages are shown in blue and green. FCs are recognized by Fasciclin 3 staining (shown in red). B. 3D reconstruction of a germarium from a multicolor lineage tracing experiment. Cells in the germarium are color-coded to illustrate the locations of FSCs in layer 1 (blue), layer 2 (pink), and layer 3 (green). Figure adapted from Reilein et al. (1).
History of FSC numbers
The number of stem cells was first calculated as the reciprocal of the fraction of the FC epithelium covered by a marked FSC clone and was measured at 9-11 days after clone induction to be sure that all transient FC clones had exited the ovariole [2]. Care was taken that the FSC clones examined likely originated from a single marked cell by inducing clones at low frequency with a heat-shock inducible recombination event, but an implicit assumption was made that FSCs divide asymmetrically (to produce one stem cell and one non-stem cell at each division) and that there would therefore be one marked FSC throughout the history of generating marked FCs. At the time, this was a common assumption for stem cells, inspired in part by studies of ovarian Germline Stem Cells. We found that FSCs do not repeatedly undergo asymmetric divisions but rather exhibit frequent amplification or loss of individual lineages (see later). An accurate count of FSCs deduced from FC contributions cannot therefore be made at 9 days after clone induction because by this time a single surviving clone will include an unknown number of FSCs. Earlier results therefore reflect the FC contribution of multiple FSCs, leading to a large underestimate of the total number of FSCs.
In 2001, Zhang and Kalderon used 3 genetic lineage labels and detected up to 3 different FSC lineage colors in a single ovariole 8 days after clone induction, concluding that there were likely three FSCs [3]. Identification of an FSC lineage was constrained by the requirement to identify an FC-contributing, surviving FSC in a location roughly consistent with the two FSC positions mapped by Margolis and Spradling; any marked FSCs that were lost prior to 8d, that occupied an unexpected position or did not contribute any FCs over the last five days would not have been scored. In 2007 Nystul and Spradling also used 3-genotype labeling and postulated 2 fixed niches of FSCs on opposite sides of the germarium, stating that the germarium is bilaterally symmetric and lies down on the slide in a particular orientation such that the two FSC niches are on either side of the germarium [4]. They did sometimes see three FC genotypes in an ovariole, but postulated that an FSC daughter could temporarily occupy one of the niches and act as an FSC.
In our more recent experiments we used the same strategies as before (counting the contribution of a single FSC lineage to the FC epithelium or counting the number of distinctly colored FSC lineages that can be generated in a single epithelium) but concluded that there are many more FSCs. The biggest reason for the very different outcome was the consideration that some FSC lineages might be lost very quickly while others amplify, necessitating looking at outcomes at a variety of times after marking. Other important differences included using more colors for lineage marking; consideration of more potential locations for FSCs and the possibility that a marked FSC may only occasionally give rise to an FC; and thorough analysis of all cells in a set of ovarioles selected without bias. We developed a lineage tracing method that yields 6 different combinations of GFP, RFP and β-galactosidase. Our strategy was to induce as many recombinations as possible in dividing stem cells in order to see the maximum number of FSC lineages. Using the enduring definition of a surviving marked FSC (a cell that remains in the 2a/b region and has given rise to at least one FC patch in the same ovariole), we found up to six (the maximum we could score) surviving FSC lineages in a single ovariole and inferred from the number of colors seen in a large sample of ovarioles that there were about five surviving FSC lineages on average.
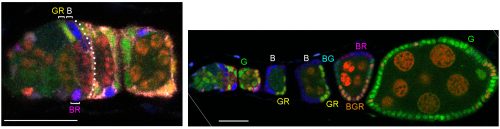
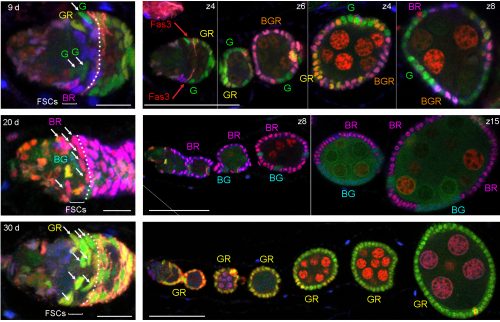
A germarium (left) and its ovariole from multicolor lineage analysis 9 days after clone induction with three FSC phenotypes (B, GR, BR) anterior to Fasciclin 3 staining (overlaid by dotted white line) in this z section. Bars, 20 microns. Image from Reilein et al. (1).
We also found that a single FSC lineage contributed to about 15% of the FC epithelium in a highly mosaic fashion with zero contribution to several egg chambers. Most important, counting surviving FSC lineages at different times after marking showed that many more lineages were present at 5d, while there were decreasing numbers at later times. We estimated that there may be about ten surviving FSC lineages at 5d, but we would need more than six color combinations to count more accurately (five or six colors were present in almost all ovarioles). Consequently, our extrapolation that there may have originally been as many as sixteen FSC lineages at day zero is also just an approximation. In these experiments we did not change the basic methodology or logic of deducing FSC numbers: every one of the cells we called an FSC remained in the germarium for longer than an FC can and had produced at least one FC, defining it unequivocally as an FSC.
Even after taking stem cell loss and amplification into account, it turns out that there is a further limitation on what was counted as a functional FSC because we required proof that an FSC produced an FC. It turns out that some of the roughly sixteen FSCs in a germarium, all of which we believe to have the potential to produce FCs, do not produce any FCs in the 4-5 day time span displayed on a fixed ovariole. This stems in essence from the apparently stochastic behavior of FSCs and positional heterogeneity within the FSC population.
FSCs are radially distributed in layers
In order to identify positions of FSCs we had to examine only those FSC clones with a single candidate stem cell (a single FSC of a certain color that matched an FC patch). This approach allowed us to define multiple locations for an FSC anywhere around the circumference of the germarium and in any of three anterior-posterior (A/P) layers adjacent to the Fasciclin 3-expressing FCs.
One of the challenges we faced as we started to realize that there were many more FSCs than expected was how to score them in sufficient detail. Eventually we developed spreadsheets to record, for each germarium and ovariole, every cell in the 2a/b region by A/P position, and the proportion of each egg chamber occupied by each color. This level of detail was essential to see where FSCs are located and to measure contributions to the FC epithelium. Tabulating the total number of cells in the 2a/b region where FSCs could be found, “the FSC region,” (16 cells on average) provided a second means to deduce the total number of FSCs. The frequency of clones with a single FSC in a given layer was very similar to the distribution of all somatic cells in the FSC region (about 8/16 in layer 1, 6/16 in layer 2 and 2/16 in layer 3, on average), suggesting that all of the roughly 16 cells are FSCs.
At the same time that we started scoring FSCs around the entire circumference, we were surprised to find by live imaging that cells in the FSC region do not stay in fixed locations but undergo back-and-forth radial movements. In addition, the ring of cells around the circumference is indistinguishable when examined by many Gal4 expression patterns and markers [5]. The existence of apparently identical interchangeable cells around the circumference of the germarium at a given A/P position suggests that each ring of FSCs constitutes a pool of equivalent cells.
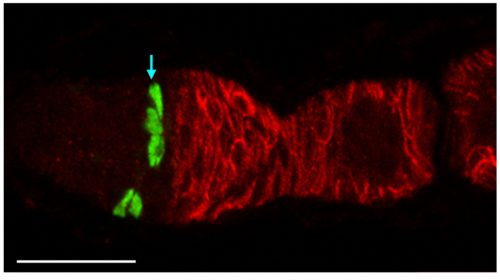
This GFP-labeled clone of FSCs (green cells indicated by arrow) illustrates how FSCs are found in multiple radial locations around the germarium. Four GFP-labeled FSCs are in layer 2 and one is in layer 3. Fasciclin 3 staining is in red. Six z-sections are combined into a maximum projection image. Bar, 20 microns. Image from Reilein et al. (1)
Population asymmetry
Fundamental to revising our ideas about FSC numbers and behavior is the realization that they are governed by population asymmetry. Population asymmetry is generally understood to mean that the daughters of a stem cell do not necessarily have different fates (one stem and one non-stem cell). Instead, within a homogeneous stem cell population of fixed size, individual stem cells undergo neutral competition during which a stem cell is frequently and stochastically duplicated or lost. Short-lived stem cells are not pre-determined or different from long-lived stem cells—they are selected by chance. If you look at the starting population you can say that only a small subset of those cells will survive as lineages for long time periods but you don’t know which ones. Hence, all of the cells are equivalent and have the potential to survive for a long time and are appropriately given the same name—stem cells. One would have to look very early after marking stem cells to capture them all before any are lost, which is often not practical. Counting the lineages after an arbitrary delay yields the number of stem cell lineages that survived for that time, and eventually only a single lineage will remain. Population asymmetry is by now a well-recognized arrangement that applies to many stem cells, including mammalian gut stem cells[6, 7]. In the mouse small intestine there are around 16 stem cells per crypt but several weeks after labeling individual cells with multiple colors only 1-3 lineages remain in most crypts[7]. We have exactly analogous findings. In both cases researchers first looked long after stem cell marking and, accordingly, thought there were a smaller number of stem cells. The appreciation of the dynamics of population asymmetry resolves these apparent contradictions in both cases.
Our data counting colored patches in ovarioles with no corresponding FSC at 9d clearly showed that many FSC lineages are lost rapidly. By 30 days the majority of ovarioles are monoclonal (we found an average of 1.5 lineages out of 60 ovarioles scored). There is a corresponding increase in the number of FSCs per lineage over time. This demonstration of rapid FSC lineage loss along with a corresponding increase in the number of 2a/b cells per surviving lineage over time shows that FSCs are maintained by population asymmetry.
Examples of germaria (3-5 z sections combined) and their ovarioles from multicolor lineage experiments 9, 20, and 30 days after clone induction. The number of distinct FSC lineages declines from four at 9 days to two (BG, BR) at 20 days to one (GR) at 30 days. The number of FSCs of one colour (white arrows) is higher when fewer lineages are present. White dotted lines indicate the borders of Fasciclin 3-expressing follicle cells. Scale bars, 10 microns (germaria) and 50 microns (ovarioles). Images from Reilein et al. (1).
Origin of Escort Cells—why had it been difficult to see the relationship to FSCs?
ECs are also labeled in lineage experiments in adults, showing that they are renewed in adult ovaries. However, Margolis and Spradling concluded that there is no lineage relationship between FSCs and ECs [2]. Kirilly et al. concurred but found that the source of new ECs is region 2a/b, very close to FSCs, postulating that the dividing cells are themselves ECs [8]. In these and other lineage studies, including ours, one could readily see ovarioles with marked FCs but no ECs, or marked ECs but no FCs, but there were also many ovarioles containing marked ECs and FCs (as well as FSCs). It was challenging to unravel the basis of these different outcomes.To prove that an FSC can give rise to both ECs and FCs it is essential to prove that a marked lineage containing all three cell types derived from a single cell. That is normally accomplished by inducing marked cells at very low frequency. We found it difficult to label single FSCs using the MARCM technique even by using the mildest of heat-shocks. It is understandable that it is quite difficult to target a single FSC for recombination by chance, given that there are about 16, rather than 2-3 per germarium. We eventually accomplished a satisfactorily low clone induction by additionally taking advantage of multicolor labeling to produce a very low frequency of clones with a specific color combination.
We also became convinced that all adult-born ECs derived from FSCs because the locations of cells in region 2a/b that proliferate, according to EdU labeling, is the same as FSC locations deduced from our multicolor labeling; it was previously thought that most of these cells were ECs. We can now categorize an EC not only by location and morphology but also by the unifying property of exhibiting no proliferation. However, it was puzzling that many lineages contained only ECs or only FCs if all of their progenitors can give rise to both ECs and FCs. We resolved this paradox by a combination of careful analysis of FSC lineages over time and the appreciation that the FSC population is heterogeneous. FSCs can produce only FCs for a few days in succession but a longer time-course shows that virtually all FC-producing lineages eventually produce ECs. Conversely, FSCs can produce only ECs for a few days; ovarioles only reveal FC production over the past 4-5 days, so a constant fraction of FSCs (around 20%) are not associated with marked FCs even though FC production may have been in their past or future. The explanation for periods of producing only ECs or only FCs lies in positional heterogeneity of FSCs.
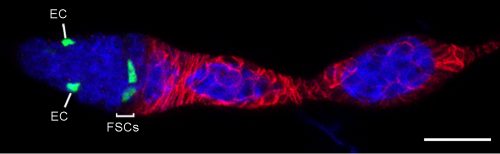
An example of a GFP-labeled FSC clone that contains only FSCs and ECs. Bar, 20 microns. Image from Reilein et al. (1).
Heterogeneity of FSCs
We found graded cell division rates and graded marker expression across FSC layers. Most important, we found that recent FC production was tightly correlated with FSC location in the most posterior layer (layer 1), leading us to postulate that FCs come directly only from layer 1 FSCs. The process of associating irreversibly with a germline cyst passing through the FSC region has never been visualized, so there was no clear prior indication of where an FC is first “specified.” ECs are anterior to FSCs, so it is almost inevitable that they must derive directly from anterior FSCs, and we also observed movement of anterior FSCs to the EC region by live imaging. Thus, the direct precursors of FCs and ECs are in different FSC layers and as long as an FSC lineage is confined to a single layer it will produce only ECs or only FCs, as often observed at short times after FSC marking. At later times, most surviving FSC lineages amplify and occupy more than one layer, followed by production of both ECs and FCs. Those FSC lineages that are then lost will leave a residue of only, generally long-lived, ECs. Movement of FSCs between layers was also inferred more directly by finding the derivatives of single marked cells in more than one layer at a reasonably high frequency even within 3 days.
The heterogeneity of FSCs presents a complication in identifying FSCs by markers because many proteins that are highly expressed in Escort cells are graded in expression across the FSC domain. For example, more anterior FSCs express much higher levels of the Wnt reporter Frizzled3-RFP and the enhancer trap PZ1444 than posterior FSCs. Other markers, such as Castor, are roughly evenly expressed across FSC layers but are also expressed in FCs. We found that only Fax-GFP stood out as being enriched in FSCs compared with ECs or FCs.
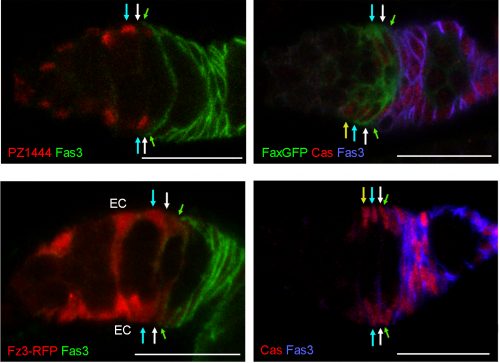
Expression of markers along the A/P axis; arrows indicate the border of Fasciclin 3 expression (green), layer 1 (white), 2 (cyan), and 3 (yellow) FSCs. PZ1444-lacZ and the Fz3-RFP Wnt pathway reporter show strong expression in ECs and graded expression across the FSC layers. Fax-GFP is expressed more strongly in FSCs than in FCs or ECs. Castor (Cas) stains FCs and all FSCs. Scale bars, 20 microns. Figure adapted from Reilein et al. (1).
Wnt signaling
We had for many years been puzzled as to why FSCs were lost at an enhanced rate when Wnt pathway activity was either eliminated or increased in FSCs. For Hedgehog and JAK-STAT pathways the results were simpler; more activity caused better survival in competition with wild-type neighbors. With our new understanding of FSC organization and behavior we could look more carefully at the consequences of manipulating Wnt pathway activity, and what we found was straightforward and striking. When Wnt pathway activity is eliminated, FSCs of that lineage become heavily biased towards occupying the most posterior FSC layer and produce FCs, with virtually no EC production. On the other hand, excess Wnt signaling forces FSCs at first into the more anterior FSC layers and eventually to become ECs. These results showed that the strength of Wnt signaling influences the A/P location and fates of FSCs.
Similarity to mammalian epithelial stem cells
With the updated organization of FSCs uncovered, we were amazed to find how similar FSC organization is to mammalian intestinal stem cells. Both FSCs and intestinal stem cells give rise to a constant supply of polarized epithelial cells. Similarities include the number of cells per niche, the organization of stem cells in heterogeneous layers, and use of Wnt as a critical niche factor[9]. Mammalian gut stem cells can produce transit amplifying (TA) or quiescent secretory cells, including Paneth cells, which eventually are retained in the crypt while TA cells migrate away. The initial location of stem cell derivatives that lead to these different outcomes are not clearly defined. Hence the spatial cues for differences in signaling pathway activities (Notch and Wnt) involved in deciding outcomes are not clear. FSCs appear to present a slightly different and simpler paradigm that may also inspire enquiry into an analogous organizational principle for gut stem cells. Wnt signaling is graded across the A/P axis of the FSC community. The relative levels of Wnt signaling dictate whether FSCs adopt more posterior positions and yield predominantly FCs (low Wnt pathway activity) or adopt more anterior positions and become ECs (high Wnt pathway activity), in accord with the natural gradient of Wnt pathway activity in this region. The paradigm of different direct differentiation outcomes for a stem cell population depending on stem cell location might apply to mammalian stem cell communities as well.
Amy Reilein and Daniel Kalderon
Department of Biological Sciences, Columbia University, New York, NY
References
Reilein, A., et al., Alternative direct stem cell derivatives defined by stem cell location and graded Wnt signalling. Nat Cell Biol, 2017. 19(5): p. 433-444.
Margolis, J. and A. Spradling, Identification and behavior of epithelial stem cells in the Drosophila ovary. Development, 1995. 121(11): p. 3797-807.
Zhang, Y. and D. Kalderon, Hedgehog acts as a somatic stem cell factor in the Drosophila ovary. Nature, 2001. 410(6828): p. 599-604.
Nystul, T. and A. Spradling, An epithelial niche in the Drosophila ovary undergoes long-range stem cell replacement. Cell Stem Cell, 2007. 1(3): p. 277-85.
Hartman, T.R., et al., Novel tools for genetic manipulation of follicle stem cells in the Drosophila ovary reveal an integrin-dependent transition from quiescence to proliferation. Genetics, 2015. 199(4): p. 935-57.
Lopez-Garcia, C., et al., Intestinal stem cell replacement follows a pattern of neutral drift. Science, 2010. 330(6005): p. 822-5.
Snippert, H.J., et al., Intestinal crypt homeostasis results from neutral competition between symmetrically dividing Lgr5 stem cells. Cell, 2010. 143(1): p. 134-44.
Kirilly, D., S. Wang, and T. Xie, Self-maintained escort cells form a germline stem cell differentiation niche. Development, 2011. 138(23): p. 5087-97.
Beumer, J. and H. Clevers, Regulation and plasticity of intestinal stem cells during homeostasis and regeneration. Development, 2016. 143(20): p. 3639-3649.
 (1 votes)
(1 votes)
 Now, on p.
Now, on p. 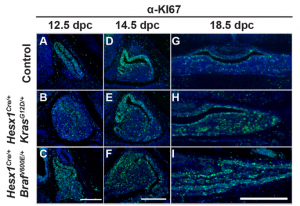 Now, on p.
Now, on p. 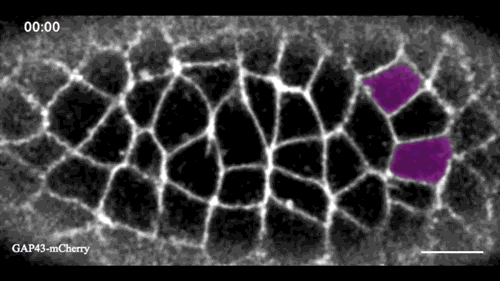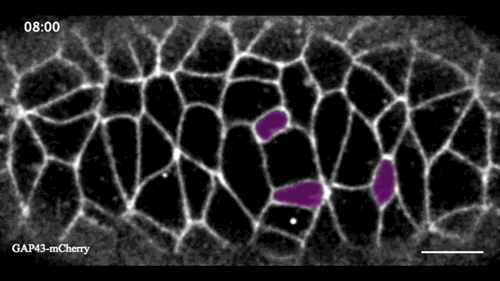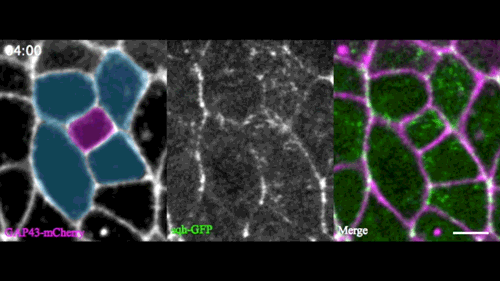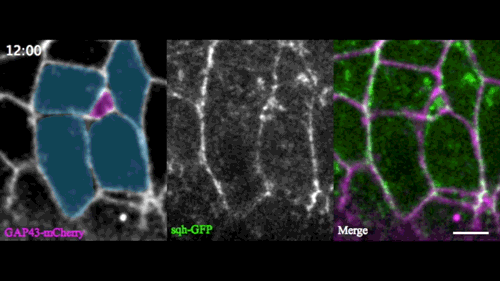
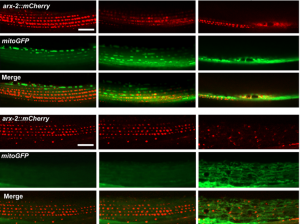 The authors show that secreted MSPds promote the localisation of mitochondria within the body wall muscle during development, and that this is dependent on signalling events that involve the CLR-1 Lar-like phosphatase receptor. Using a targeted RNAi screen of known genes implicated in ALS and SMA, the authors identified survival of motor neuron 1 (SMN-1) as a crucial downstream mediator of MSPd and further showed that SMN-1 and ARX-2 are important for mitochondria localisation along the I-bands in body wall muscle.
The authors show that secreted MSPds promote the localisation of mitochondria within the body wall muscle during development, and that this is dependent on signalling events that involve the CLR-1 Lar-like phosphatase receptor. Using a targeted RNAi screen of known genes implicated in ALS and SMA, the authors identified survival of motor neuron 1 (SMN-1) as a crucial downstream mediator of MSPd and further showed that SMN-1 and ARX-2 are important for mitochondria localisation along the I-bands in body wall muscle.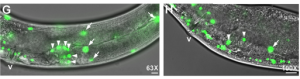 The authors further demonstrate that, under normal conditions, gonad development depends on germline and neuronal expression of vpr-1 and specifically on cleaved MSPd, which circulates in the pseudocoelom and induces gonadogenesis non-cell-autonomously, much like a hormone.
The authors further demonstrate that, under normal conditions, gonad development depends on germline and neuronal expression of vpr-1 and specifically on cleaved MSPd, which circulates in the pseudocoelom and induces gonadogenesis non-cell-autonomously, much like a hormone. (No Ratings Yet)
(No Ratings Yet)