DMM is looking for an enthusiastic intern who wishes to gain experience in science publishing, including writing press releases, contributing to our social media activities, and supporting our Reviews Editor with commissioned articles. The internship is envisaged to last for up to 1 year at a salary of £15,000 p.a.
Our interns have a great track record of continuing on into important publishing roles.
Joining an experienced and successful team, the internship offers an ideal opportunity to gain in-depth experience on a growing Open Access journal in the exciting and fast-moving field of translational research. DMM publishes primary research articles and a well-regarded front section, including commissioned reviews and poster articles, thought-provoking editorials and interviews with leaders in the field. We also have an active social media presence and will be growing our press release programme. The intern will work alongside an established publishing team in our Cambridge offices.
Because the journal serves both basic biomedical researchers and clinicians, applicants will have a PhD or MD, ideally with some relevant research experience, and a broad knowledge of model organisms and disease issues.
Is the role for you? You would be expected to…
Support our Reviews Editor:
• Identify and commission topical front-section content from top-ranking scientists, see articles through peer review and work closely with authors to finalise articles for publication.
• Travel to international scientific conferences and research institutes, representing the journal, keeping abreast of the latest research and making contacts in the DMM community.
Develop your own areas of activity:
• Spot newsworthy articles, write informative press releases and handle any media enquiries.
• Interview high-profile scientists in the biomedical arena.
• Contribute to our social media output.
• Be creative – contribute other ideas for the journal’s development and promotion.
Essential requirements for the job are enthusiasm, commitment, judgement and integrity. Candidates should have excellent interpersonal skills and confidence, excellent oral and written communication skills, and a broad interest in research and the research community. They should also be willing to travel. Previous editorial experience is not required, but we would expect candidates to be able to demonstrate an interest in scientific communication.
A post-doctoral position is available in the Franz-Odendaal Bone Development Lab to study the developmental basis of the vertebrate ocular skeleton in a comparative context. Highly motivated and independent individuals with excellent interpersonal skills are encouraged to apply. The successful applicant will take a key role in our research program which interests spans evo-devo, developmental genetics and phenotypic variation.
We are seeking a recent doctoral student in Biological Sciences or related fields with experience in Molecular Biology, Cell and Developmental Biology. Experience with zebrafish and/or chick embryos is desirable but not required. Opportunities to supervise students will be available.
Please send curriculum vitae and summary of research interests via email.
Dr Tamara Franz-Odendaal
Franz-Odendaal Bone Development Lab
Mount Saint Vincent University,
Halifax, Nova Scotia, Canada
We are offering a Research Assistant position funded by a Wellcome Investigator Award. The successful candidate for this position will perform experiments and provide technical support in the group of James Briscoe. The group studies the molecular and cellular mechanisms responsible for the embryonic development of the vertebrate nervous system.
performing research projects involving molecular biology, tissue culture and embryological techniques
providing technical advice and teaching basic techniques to new members
interacting with colleagues by discussing results and techniques and ideas for improvements
overseeing the effective running of the lab by monitoring stock levels and ordering consumables and reagents, maintaining equipment, databases and methods sheets.
The Francis Crick Institute (the Crick) is a registered charity whose purpose is to conduct biomedical research into all aspects of human health and disease. Dedicated to research excellence, the institute will be a world-leading centre of biomedical research and innovation. The Crick is located in a new, purpose-built research centre in central London (next to St Pancras International), housing 1,250 researchers and 250 support staff.
The new Program in Functional and Chemical Genomics at the Oklahoma Medical Research Foundation (OMRF, http://omrf.org/) is inviting applications for multiple independent faculty positions at the Assistant and Associate Member levels (Assistant or Associate Professor equivalent). OMRF is a non-profit private foundation dedicated to fundamental, interdisciplinary research and to the translation of knowledge to medicine and public health in areas including autoimmune disease, cardiovascular disease, aging-related diseases, metabolic diseases and cancer. The Program will combine the latest technologies in genomics, functional genomics, model systems biology, chemical biology and computational analyses to tackle fundamental questions emerging from ongoing high-throughput genetic and epigenetic studies of disease. The Program occupies newly renovated space designed to foster highly collaborative and innovative research using computational and/or genetic models systems such as yeast, C. elegans, Drosophila, zebrafish, mice and stem cells aimed at scaling the evaluation of genetic and epigenetic alterations. This biological understanding will be used to interrogate opportunities for therapeutic intervention and define chemical mechanisms. Faculty research programs at OMRF are supported by competitive start-up and salary packages, as well as, ongoing annual support and a commitment to faculty development. OMRF faculty have access to state-of-the-art mouse and zebrafish facilities and outstanding core technology laboratories like Flow Cytometry, Imaging, NextGen Sequencing and more.
We are looking for interactive and creative faculty whose research interests synergize with, but expand, the Foundation’s current research expertise. Candidates holding a Ph.D. and/or M.D. in biological or biomedical science fields, with relevant postdoctoral experience, and outstanding research accomplishments in areas including, but not limited to the following are invited to apply immediately:
• Functional Genomics using Genetic Model Systems
• Drug Target Validation and Chemical Screening
• Stem and Progenitor Cell Biology
• Cell Fating and Developmental Biology
• Genetics and Epigenetics Interactions
• Systems Biology
Successful candidates are expected to develop and maintain robust extramurally funded research programs and may seek adjunct appointments in one of the many relevant University of Oklahoma School of Medicine and/or Graduate College Departments, and will have opportunities to participate in one or more graduate student education programs.
To apply, submit the following documents via FunctionalChemicalGenomics@omrf.org : 1) Cover Letter; 2) Curriculum Vitae; 3) Research Statement; 4) Contact information for 3 references.
We are seeking outstanding candidates to lead a project studying Notch, TGF-β, and ephrinB2 signaling pathways in arterial venous programming/reprogramming and the implication in development and diseases. We take a conditional mouse genetic approach to manipulating gene expression in endothelial cell-specific and temporally controlled fashion. We also use cutting-edge in vivo real time imaging technology, including an in-lab constructed two-photon microscope, which provides exceptional access to gene function in vivo at the cellular resolution along with blood flow measurement overtime in live animals. This basic approach is complemented by preclinical studies with our elegant mouse models of diseases, offering outstanding opportunities for translational research. The laboratory is well equipped with state-of-the-art capabilities at the molecular, cellular, and organismic levels. In addition to funding from the PI, we also have an excellent track record in sponsoring postdoc fellowships. We are interested in a well-trained and well-published recent Ph.D. graduates to continue our innovative breakthroughs in a rewarding training program. This postdoctoral research is an excellent platform for a highly productive Ph.D. with a strong motivation to become a future group leader. Experience with mouse techniques is a plus. UCSF offers outstanding postdoctoral career development opportunities. Please submit your CV, research interests, and the names of three references by email with a subject title “postdoc application” to:
Rong Wang, Ph. D.
Professor
UCSF
rongwangucsf@gmail.com
For additional information visit:
http://profiles.ucsf.edu/rong.wang
http://wanglab.surgery.ucsf.edu
https://bms.ucsf.edu/directory/faculty/rong-wang-phd
Each of us has around 6 pints of blood. The blood contains a number of different types of cells, including oxygen-transporting red blood cells, disease-protecting white blood cells or wound-closing platelets. But have you ever wondered where they all come from?
Quite amazingly, all these very different blood cells originate from the same parental cell, called the haematopoietic stem cell (HSC for short). HSCs live inside our bone marrow and keep making new blood cells throughout life. That’s why you don’t have to worry if you cut yourself and lose some blood – your bone marrow will make new cells very quickly. In fact, a single haematopoietic stem cell has the potential to make all 6 pints of your blood!
As it turns out, the way we make the first HSCs is very similar to all other vertebrates studied so far (Ciau-Uitz et al, 2014): they come from endothelial cells, the cells lining the vessels of the circulatory system. But only a specialised type of endothelium gives rise to HSCs – the haemogenic endothelium, located in the main artery of the 6-week old human embryo. 100 years ago, Emmel had observed blood cells associated with arterial endothelium in pig embryos (Fig.1).
Figure 1 – Transversal section of the main artery (the dorsal aorta) of a pig embryo. Adapted from Emmel, 1916.
At that time, similar observations were made in a miriad of other vertebrate embryos, including the mongoose, the chick, the rabbit and the human (see Adamo and Garcia-Cardena, 2012 for the full references). This abundance of early observations led to the hypothesis that blood cells came from… blood vessels! The evidence to support this very simple hypothesis didn’t come until 2010, when a few research groups imaged the birth of an HSC in live zebrafish embryos (Bertrand et al., 2010; Kissa and Herbomel, 2010; Lam et al., 2010).
In the Patient lab, we use zebrafish to find out what makes these endothelial cells, already part of a differentiated tissue, become our all-important HSCs. In our recent Developmental Cell paper, we solved another piece of the puzzle: we showed that the cytokine Transforming Growth Factor β (TGFβ) is needed very early in the developing embryos to make the endothelium become haemogenic, so that it can make HSCs. Here is the story of how we got there.
Why TGFβ?
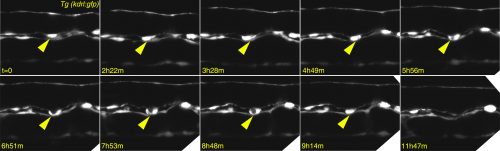
Any haematologist will tell you that if you give TGFβ to adult mice, their blood stem cells will stop proliferating and, if exposed for too long, they will die. So why would you need TGFβ to make the stem cells in the first place and why on Earth would you bother to even look at TGFβ? Well, the clue comes from epithelial cancers: an oncologist will tell you that TGFβ is a tumour suppressor that keeps cancer cells from proliferating and even from moving to other parts of the body…until it doesn’t! After a fatal tipping point, TGFβ becomes an oncogene and actually encourages the cancer cells to transform and go invade other tissues. This is what led us to look at TGFβ in the formation of HSCs – an endothelial cell leaving its place in a functional vessel and becoming an HSC (see Figure 2 for our own live imaging of the birth of an HSC) is remarkably similar to a metastasizing cancer cell!
Figure 2 – Confocal timelapse imaging of an emerging haematopoietic stem cell in the zebrafish dorsal aorta (yellow arrow)
This was a great opportunity to put together my interest in developmental biology and in stem cells, and hopefully contribute with new discoveries that may also be relevant to human biology. The first signs were encouraging: the TGFβ receptor tgfβR2 and its ligands tgfβ1a and tgfβ1b were present in the main embryonic artery at the right time; we also found tgfβ2 and tgfβ3 in the neighbouring notochord, further suggesting TGFβ signalling might play a role. Knocking down the receptor indeed led to the loss of haematopoietic cells, so we were in business! I convinced the British Heart Foundation that TGFβ was a good idea and got an Intermediate Fellowship to go ahead with this line of research, hosted in the Patient lab at the MRC Weatherall Institute of Molecular Medicine. You can find out more about other research ongoing at the MRC WIMM on the WIMM blog.
The first pieces of the puzzle
After we demonstrated that TGFβ was required to make the HSCs, we wanted to figure out how it related to other pathways known to play a role in the process. We turned to NanoString technology, a neat multiplex hybridization technology to look at what happened to gene expression downstream of TGFβ. Of the 100 plus genes we looked at, one turned out to be activated by TGFβ – the Notch ligand jag1a. This was a crucial finding, because jag1 had already been described as a target of TGFβ in metastasizing cancer cells (Zavadil et al., 2004) and so the link to Notch signalling was already established. We followed this lead and discovered that switching off jag1a also resulted in losing HSCs. Moreover, forced expression of jag1a rescued the loss of HSCs in TGFβ-deficient embryos, further supporting the TGFβ-Notch link.
Through a series of experiments that involved switching off other important cell signalling pathways and subsequent gene expression analysis, we managed to place the TGFβ pathway downstream of VEGF – a signal that is a well known player in the development of blood and blood vessels.
The final piece
At this point, we were excited that we had a nice VEGF-TGFβ-Notch story to tell, but we were not finished yet! We wanted to see which of the TGFβ ligands was doing the job. There are a number of different family members (TGFβ1, TGFβ2, TGFβ3), all of which trigger very similar events in their target cells. To our surprise, not only our prime suspects (based on expression analysis), TGFβ1a and b were required, but also TGFβ3 played a role. Even more surprisingly, TGFβ3 was more important at a later stage, when the HSCs actually leave the endothelium and become motile. Sticking to the similarities with cancer, this would be when the cancer cells metastasize. In short, while TGFβ1 and TGFβ3 were required to make the HSCs, they came from different sources and were required at different times. This finding really puts emphasis on how important it is to consider the timing of events when studying embryonic development.
Is this the end of the story?
No!… This is really just the beginning. What we discovered is that there is a ‘window of opportunity’ where TGFβ is required, but we and others have shown that if you have too much TGFβ you will also struggle to make HSCs (Nimmo et al., 2013; Vargel et al., 2016; Yang et al., 2016). Think of this like you’re cooking a meal: when you add salt, too much or too little of it will ruin your dish, so it’s important to get it right. How can we reconcile both observations? Also, how does TGFβ3 ‘take over’ the role of the main TGFβ ligand and how is it regulated? This is one of the most exciting times in the life of a researcher – when you get an answer that comes with… many more questions!
How can understanding the origins of our blood be useful in the long term? If we discover enough pieces of the puzzle, we may be able to write down a trusted ‘recipe’ to prepare the haematopoietic stem cells in a laboratory dish, step by step. Such optimised, lab-grown HSCs would have a great potential to help people suffering from various blood disorders, including leukaemias. Let’s hope we’ll be able to start ‘cooking’ soon!
Contributors: Rui Monteiro and Tomasz Dobrzycki
References
Adamo, L., and Garcia-Cardena, G. (2012). The vascular origin of hematopoietic cells. Dev Biol 362, 1-10.
Bertrand, J.Y., Chi, N.C., Santoso, B., Teng, S., Stainier, D.Y., and Traver, D. (2010). Haematopoietic stem cells derive directly from aortic endothelium during development. Nature 464, 108-111.
Kissa, K., and Herbomel, P. (2010). Blood stem cells emerge from aortic endothelium by a novel type of cell transition. Nature 464, 112-115.
Lam, E.Y., Hall, C.J., Crosier, P.S., Crosier, K.E., and Flores, M.V. (2010). Live imaging of Runx1 expression in the dorsal aorta tracks the emergence of blood progenitors from endothelial cells. Blood 116, 909-914.
Nimmo, R., Ciau-Uitz, A., Ruiz-Herguido, C., Soneji, S., Bigas, A., Patient, R., and Enver, T. (2013). MiR-142-3p controls the specification of definitive hemangioblasts during ontogeny. Dev Cell 26, 237-249.
Vargel, Ö., Zhang, Y., Kosim, K., Ganter, K., Foehr, S., Mardenborough, Y., Shvartsman, M., Enright, A.J., Krijgsveld, J., and Lancrin, C. (2016). Activation of the TGFβ pathway impairs endothelial to haematopoietic transition. Scientific reports 6, 21518.
Yang, Q., Liu, X., Zhou, T., Cook, J., Nguyen, K., and Bai, X. (2016). RNA polymerase II pausing differentially regulates signaling pathway genes to control hematopoietic stem cell emergence in zebrafish. Blood.
Zavadil, J., Cermak, L., Soto-Nieves, N., and Bottinger, E.P. (2004). Integration of TGF-beta/Smad and Jagged1/Notch signalling in epithelial-to-mesenchymal transition. EMBO J 23, 1155-1165.
The current issue of Development – our ‘Special Issue on Plant Development’ – contains a collection of review- and research-based articles focusing on plant development.
Below, you can find details of the review-based articles in this Special Issue:
Plant development: a Special Issue
Ottoline Leyser introduces this Special Issue focusing on plant developmental biology, which is published in honour of Ian Sussex – a founding father of the field. Read the Editorial on p. 3223
Ian Sussex: simple tools, clever experiments and new insights into plant development
Scott Poethig looks back at the research career of Ian Sussex, who helped transform the discipline of plant developmental biology into the dynamic, sophisticated field that it is today. Read the Spotlight article on p. 3224
Mechanisms of auxin signaling
The plant hormone auxin triggers complex growth and developmental processes. In their poster article, Meirav Lavy andMark Estelle provide an overview of the auxin signal transduction pathway, highlighting how it facilitates rapid switching between transcriptional repression and gene activation. See the Development at a Glance article on p. 3226
The Sussex signal: insights into leaf dorsiventrality
The differentiation of a leaf – from its inception as a semicircular bulge on the surface of the shoot apical meristem into a flattened structure with specialized upper and lower surfaces – is one of the most intensely studied processes in plant developmental biology. Here, Cris Kuhlemeier andMarja Timmermans revisit Ian Sussex’s early studies of leaf dorsiventrality and describe our current understanding of the mechanisms that establish and maintain adaxial-abaxial leaf polarity. See the Review on p. 3230
CLAVATA-WUSCHEL signaling in the shoot meristem
Shoot meristems are maintained by pluripotent stem cells that are controlled by CLAVATA-WUSCHEL feedback signaling. This pathway, which coordinates stem cell proliferation with differentiation, was first identified in Arabidopsis, but appears to be conserved in diverse higher plant species. Here, Rüdiger Simon, David Jackson and colleagues highlight the commonalities and differences between CLAVATA-WUSCHEL pathways in different plant species. See the Review on p. 3238
Developing a ‘thick skin’: a paradoxical role for mechanical tension in maintaining epidermal integrity?
Plant aerial epidermal tissues, like animal epithelia, act as load-bearing layers and play pivotal roles in development. The presence of tension in the epidermis has implications for organ shapes but it also constantly threatens the integrity of this tissue. Here, Olivier Hamant, Gwyneth Ingram and co-workers explore the relationship between tension and cell adhesion in the plant epidermis, and examine how tensile stress perception may act as a regulatory input to preserve epidermal tissue integrity and thus normal morphogenesis. See the Review on p. 3249
MADS-domain transcription factors and the floral quartet model of flower development: linking plant development and evolution
The floral quartet model of floral organ specification poses that different tetramers of MIKC-type MADS-domain transcription factors control gene expression and hence the identity of floral organs during development. Here, Günter Theißen and colleagues provide a brief history of the floral quartet model and review several lines of recent evidence that support the model. They also suggest a novel hypothesis describing how floral quartet-like complexes may interact with chromatin during target gene activation and repression. See he Review on p. 3259
Helical growth in plant organs: mechanisms and significance
Many plants show some form of helical growth, such as the circular searching movements of growing stems and other organs (circumnutation), tendril coiling, leaf and bud reversal (resupination), petal arrangement (contortion) and leaf blade twisting. Here, David Smyth provides an overview of the genes and cellular processes that underlie helical patterning, and discusses the diversity of helical growth patterns in plants, highlighting their potential adaptive significance. See the Review on p. 3732
Enhancing crop yield by optimizing plant developmental features
A number of plant features and traits, such as overall plant architecture, leaf structure and morphological features, vascular architecture and flowering time, are important determinants of photosynthetic efficiency and hence the overall performance of crop plants. The optimization of such developmental traits thus has great potential to increase biomass and crop yield. Here, Aashish Ranjan and colleagues provide a comprehensive review of these developmental traits in crop plants, summarizing their genetic regulation and highlighting the potential of manipulating these traits for crop improvement. See the Review on p. 3283
PLUS:
This Special Issue also contains a number of Research Reports, Research Articles and Techniques & Resources Articles – click here for a full listing!
Applications are invited for a Research Assistant/Associate within the Division of Infection & Immunity at UCL in Dr Gillian Tomlinson’s laboratory.
We are seeking highly motivated individuals interested in integrating cutting-edge human and zebrafish models to study the immunopathogenesis of tuberculosis. The post is funded by a Medical Research Council Clinician Scientist Fellowship entitled “Tuning the immune response in tuberculosis”, and combines a human experimental tuberculosis challenge model with studies using Mycobacterium marinum infection of zebrafish to identify and validate host factors that calibrate a favourable immune response in tuberculosis.
The post-holder will be supervised by Dr Gillian Tomlinson based in the Cruciform Building at UCL. Dr Tomlinson works with Dr Mahdad Noursadeghi’s and Professor Benny Chain’s group’s which study host immune responses to infectious diseases at genome wide level with a particular focus on tuberculosis (www.innate2adaptive.com). The zebrafish work will be supported by the fully managed world class research aquarium at UCL.
The post is available for 3 years in the first instance, subject to satisfactory probationary and annual appraisals. There is an established track record for department post-doctoral staff gaining personal fellowships. Independently minded and talented investigators will be encouraged and supported in seeking such fellowship support.
Key Requirements
Applicants must have an MSc (or equivalent degree) and/or a PhD (or equivalent degree) or about to be awarded in a relevant subject.
Candidates must have experience of working with zebrafish including microinjection and in vivo imaging.
Evidence of significant scientific contribution including publications and presentations at conferences is also essential.
Appointment at Grade 7 is dependent on having been awarded a PhD. If this is not the case, initial appointment will be at research assistant Grade 6B (salary £29,485 – £31,091 per annum) with payment at Grade 7 being backdated to the date of final submission of the PhD thesis.
To apply for this post please follow this link to the UCL website: https://atsv7.wcn.co.uk/search_engine/jobs.cgi?owner=5041178&ownertype=fair&jcode=1580418
The Department of Biology invites applications for a tenure-track position at the assistant professor level, beginning Fall 2017. Applicants should have a Ph.D.; post-doctoral experience is desirable. The strongest candidates will demonstrate a commitment to teaching and an active research program that can involve motivated undergraduates from diverse backgrounds. We seek a broadly trained systems biologist whose research and course offerings complement those of other faculty in the department. We invite applicants who use contemporary methodologies, including computational and/or modeling techniques, to study biological systems at the cellular or organismal level. Teaching responsibilities include participation in a team-taught introductory course, a one-semester intermediate level course in systems biology with a laboratory, and an advanced seminar with research projects in the area of the applicant’s interest. Participation in the interdisciplinary Neuroscience program is possible. The College offers competitive start-up packages to support faculty research and teaching, as well as a generous sabbatical policy (http://www.swarthmore.edu/provosts-office/sabbatical-support).
Located in the immediate suburbs of Philadelphia and just 20 miles from Wilmington DE, Swarthmore College is a highly selective liberal arts college whose mission combines academic rigor with social responsibility. Swarthmore has a strong institutional commitment to diversity, and actively seeks and welcomes applications from candidates with exceptional qualifications, particularly those with demonstrable commitments to a more inclusive society and world. Applicants from traditionally underrepresented groups are strongly encouraged to apply. For more information on Faculty Diversity and Excellence at Swarthmore, see http://www.swarthmore.edu/faculty-diversity-excellence/information- candidates-new-faculty.
All application materials should be submitted on Academic Jobs Online [https://academicjobsonline.org/ajo/jobs/7479] by October 8, 2016. Questions regarding this position should be addressed to the Search Chair at biosearch@swarthmore.edu.
Monash University is seeking an outstanding candidate to manage and lead the Department of Anatomy and Developmental Biology within the Faculty of Medicine Nursing and Health Sciences. The Department of Anatomy and Developmental Biology is one of six departments of the School of Biomedical Sciences and a leader within its discipline. Areas of research expertise include renal and lung biology, epithelial and reproductive biology, inflammation, embryology, cancer, stem cell biology and regenerative medicine.
The successful candidate will capitalise and build on existing educational and research strengths to create a high-performing team of academics and professional staff who not only provide outstanding educational opportunities to the students in the school but also, through membership of the Monash Biomedicine Discovery Institute, contribute to the research strategy and output of the institute.
To be successful in this role you will have a proven track record of individual research excellence and evidence of leadership through building and developing capability in an academic or research environment.
 (No Ratings Yet)
(No Ratings Yet)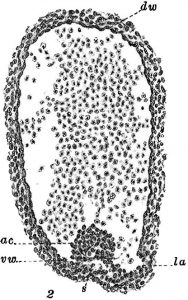
 (2 votes)
(2 votes)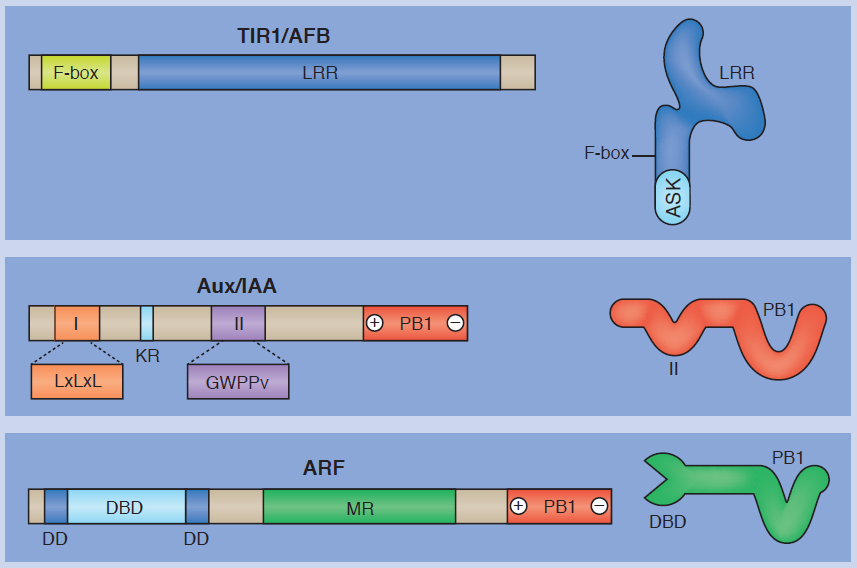 growth and developmental processes. In their poster article,
growth and developmental processes. In their poster article,  The differentiation of a leaf – from its inception as a semicircular bulge on the surface of the shoot apical meristem into a flattened structure with specialized upper and lower surfaces – is one of the most intensely studied processes in plant developmental biology. Here,
The differentiation of a leaf – from its inception as a semicircular bulge on the surface of the shoot apical meristem into a flattened structure with specialized upper and lower surfaces – is one of the most intensely studied processes in plant developmental biology. Here,  cells that are controlled by CLAVATA-WUSCHEL feedback signaling. This pathway, which coordinates stem cell proliferation with differentiation, was first identified in Arabidopsis, but appears to be conserved in diverse higher plant species. Here,
cells that are controlled by CLAVATA-WUSCHEL feedback signaling. This pathway, which coordinates stem cell proliferation with differentiation, was first identified in Arabidopsis, but appears to be conserved in diverse higher plant species. Here, 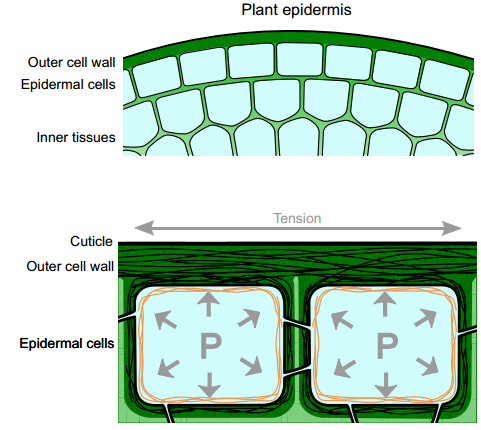 Plant aerial epidermal tissues, like animal epithelia, act as load-bearing layers and play pivotal roles in development. The presence of tension in the epidermis has implications for organ shapes but it also constantly threatens the integrity of this tissue. Here,
Plant aerial epidermal tissues, like animal epithelia, act as load-bearing layers and play pivotal roles in development. The presence of tension in the epidermis has implications for organ shapes but it also constantly threatens the integrity of this tissue. Here, 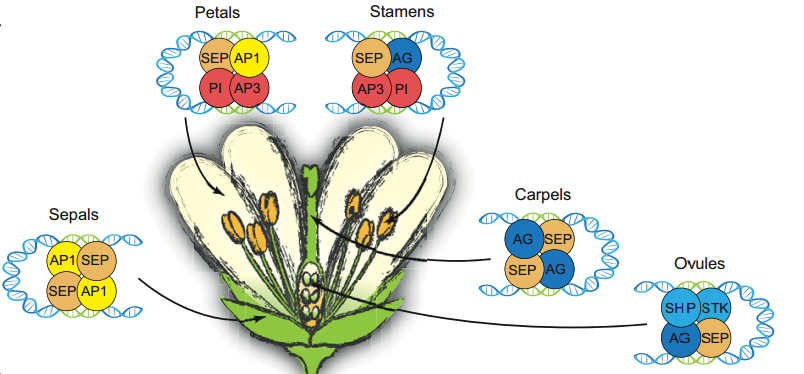 The floral quartet model of floral organ specification poses that different tetramers of MIKC-type MADS-domain transcription factors control gene expression and hence the identity of floral organs during development. Here, Günter Theißen and colleagues provide a brief history of the floral quartet model and review several lines of recent evidence that support the model. They also suggest a novel hypothesis describing how floral quartet-like complexes may interact with chromatin during target gene activation and repression. See he Review on p.
The floral quartet model of floral organ specification poses that different tetramers of MIKC-type MADS-domain transcription factors control gene expression and hence the identity of floral organs during development. Here, Günter Theißen and colleagues provide a brief history of the floral quartet model and review several lines of recent evidence that support the model. They also suggest a novel hypothesis describing how floral quartet-like complexes may interact with chromatin during target gene activation and repression. See he Review on p. 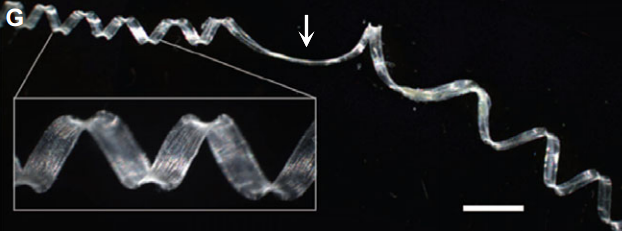 Many plants show some form of helical growth, such as the circular searching movements of growing stems and other organs (circumnutation), tendril coiling, leaf and bud reversal (resupination), petal arrangement (contortion) and leaf blade twisting. Here, David Smyth provides an overview of the genes and cellular processes that underlie helical patterning, and discusses the diversity of helical growth patterns in plants, highlighting their potential adaptive significance. See the Review on p.
Many plants show some form of helical growth, such as the circular searching movements of growing stems and other organs (circumnutation), tendril coiling, leaf and bud reversal (resupination), petal arrangement (contortion) and leaf blade twisting. Here, David Smyth provides an overview of the genes and cellular processes that underlie helical patterning, and discusses the diversity of helical growth patterns in plants, highlighting their potential adaptive significance. See the Review on p. 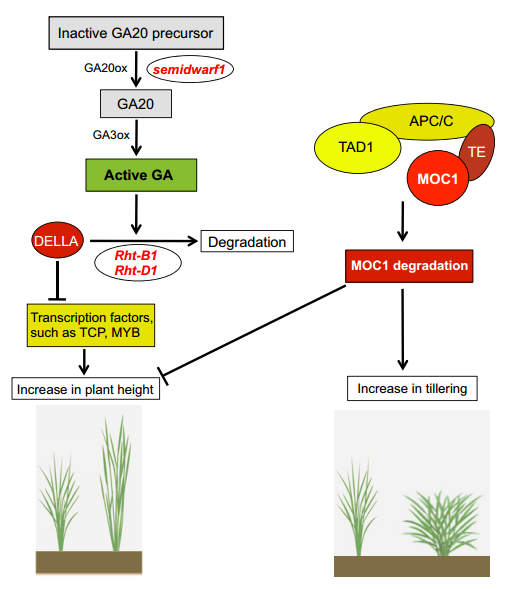 as overall plant architecture, leaf structure and morphological features, vascular architecture and flowering time, are important determinants of photosynthetic efficiency and hence the overall performance of crop plants. The optimization of such developmental traits thus has great potential to increase biomass and crop yield. Here, Aashish Ranjan and colleagues provide a comprehensive review of these developmental traits in crop plants, summarizing their genetic regulation and highlighting the potential of manipulating these traits for crop improvement. See the Review on p.
as overall plant architecture, leaf structure and morphological features, vascular architecture and flowering time, are important determinants of photosynthetic efficiency and hence the overall performance of crop plants. The optimization of such developmental traits thus has great potential to increase biomass and crop yield. Here, Aashish Ranjan and colleagues provide a comprehensive review of these developmental traits in crop plants, summarizing their genetic regulation and highlighting the potential of manipulating these traits for crop improvement. See the Review on p.