On the Air: Bringing developmental biology to the radio
Posted by Simon Bishop, on 5 February 2014
I had just put the finishing touches to my thesis, a weighty tome of developmental neurobiology. My mind was still focused on this highly specific piece of work. I was unprepared, therefore, for the first question:
“What is infinity?”
I was live on air. I couldn’t see the caller or — since I was in a studio 65 miles away — the show presenter. I had no idea how to even start to answer such a question, not least because I did not know the answer. But this was the BBC Radio Norfolk science phone-in with the Naked Scientists, a fortnightly feature, and dead air was not an option. So I started to talk — nonsense at first, but eventually finding a voice among the tangle of complex mathematics and philosophy that I had forged. It was only day one, but my internship with the Naked Scientists had truly begun.
Based at the University of Cambridge, the Naked Scientists are a group of science broadcasters with expertise covering the life and applied sciences. Their flagship programme, a one hour live programme on Sunday evenings from the studios of BBC Radio Cambridgeshire, is repeated on ABC Radio National (Australia), re-broadcast in an edited form on BBC Radio 5Live (UK national) and released as a podcast globally. In addition, the team produce a number of subject-specific podcasts — including genetics, neuroscience and astronomy — and a number of smaller features, such as the phone-ins to Norfolk and South Africa. Combined, they have a wide audience.
Having met the lead Naked Scientist, Dr Chris Smith, at a science communication workshop early last year, I managed to secure funding from the Genetics Society to join the team for two months. In that time I contributed live and pre-recorded content to a number of stations, and produced a documentary on genetics. I booked guests, conducted interviews and edited features; all of this with no prior experience in radio, just an interest in communicating science.
We all get carried away with the subjects we are passionate about: it is the subjects in which we are not an expert that allow us to better relate to audiences. I was told with two hours’ notice, for example, that I was to go on air to talk about special relativity. I was tasked with stripping down a complex and beguiling topic in three minutes of air time. I had to find a way to explain it with no prior knowledge, just like those listening. This is the very same frame of mind we should all apply to our own research areas when the opportunity for outreach arises.
But what is that audience? For the Naked Scientists, there is a unique challenge presented by its format: the programme initially airs on local radio, to an interested but non-specialist public, but is re-released online to a podcast audience that has sought scientific conversation. As such, the programmes must be approachable and detail must be built up sequentially and clearly for the first audience, but detail must not be skimped for the second. This presents a unique challenge, requiring forethought of the mix of topics included, careful vetting of guests to determine how confidently they can describe their work (and how they respond to an interviewer pretending to know nothing), and a mix of media — different voices, a mixture of live and pre-recorded content, all carefully timed.
A further challenge is, quite simply, that this is radio. Clarity of description is paramount, for diagrams and animations are invisible to the audience, as are flappy hands. On one occasion, I found myself describing what I could see on a computer screen, a curious sensation absent from normal conversation, for the benefit of listeners at home.
The first of the Naked Scientists audiences — as with all television and radio science output — means that detailed and complex topics might often be vetoed. Yet its second audience provides an out. Thus, when I was entrusted with one whole hour of live radio, to do the research, book the guests, write the script and co-present, I chose developmental biology.
Gastrulation and neurulation, the early steps, stages and movements of embryogenesis, are as perplexing to many as special relativity. To fill an hour long show with talk of these pivotal stages of development, in addition to the genetics of the actin cytoskeleton, which controls cell shape; how cells know where they are in the body; and gross morphological development, using the example of limb development and club feet, was a challenge I relished. I cannot claim to have been successful; this must be left to the audience to judge. Yet I am proud of my attempts, and very grateful to the many contributors, including Richard Adams from the University of Cambridge, Helen Matthews at University College London, Neil Vargesson at the University of Aberdeen and Max Heiman, Harvard Medical School. The programme, Super-shape me! is available from the Naked Scientists website.
I learnt much over my short time on the radio. I recommend trying radio as a medium for truly testing science outreach skills — try contacting your local student radio station or one of the many science podcast projects available. But perhaps the skill I would flag for now to readers of The Node is that of interviewing. Researchers with an interest in communication try, given the opportunity, to explain their work. But let’s turn that on its head: first, pick someone unaccustomed to outreach, whose work is on a topic you are entirely unfamiliar with. Second, pretend you know nothing about the topic or how science works. Try to get them to talk for 3, 4 even 7 minutes (radio features are, after all, of variable lengths) on a level the public could follow. What are they doing? How are they doing it? WHY are they doing it? Keeping control of such a conversation, particularly to control the jargon, is a challenge indeed. Give it a try!
![]() This post is part of a series on science outreach. You can read the introduction to the series here and read other posts in this series here.
This post is part of a series on science outreach. You can read the introduction to the series here and read other posts in this series here.
 (5 votes)
(5 votes) I think that one of the scariest parts of being President of the Society for Developmental Biology is coming up with topics for these editorials in the Newsletter.
I think that one of the scariest parts of being President of the Society for Developmental Biology is coming up with topics for these editorials in the Newsletter. (25 votes)
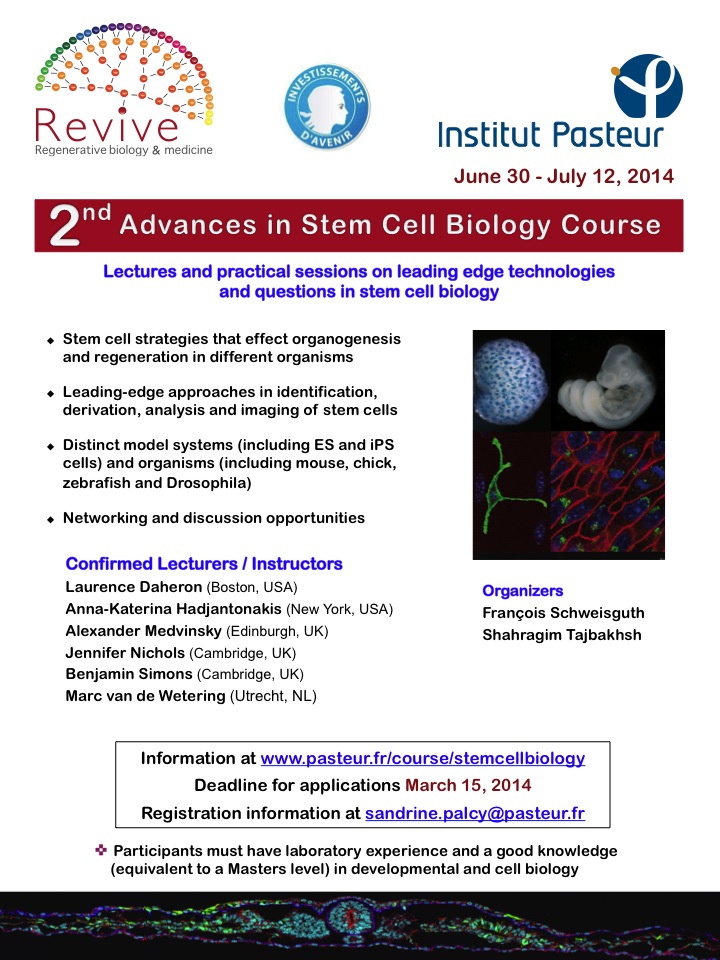
(25 votes) Within the developing neocortex, multiple progenitor cell types contribute to neuronal production, and the properties and relative abundance of these populations help to define the extent of neocortical growth. As well as apically localised radial glial cells (RGCs) that comprise the stem cell compartment, various populations of basal progenitors (BPs) exist – including basal RGCs, with both self-renewal and proliferative capacity, and more restricted progenitors. The stem cell-like properties of RGCs are linked to the fact that these cells are connected to the basal lamina, from which they receive proliferative signals via integrins. Now, Denise Stenzel et al. investigate in rodents the role of integrin αvβ3 in regulating the proliferative capacity of BPs (
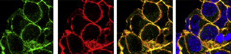
Within the developing neocortex, multiple progenitor cell types contribute to neuronal production, and the properties and relative abundance of these populations help to define the extent of neocortical growth. As well as apically localised radial glial cells (RGCs) that comprise the stem cell compartment, various populations of basal progenitors (BPs) exist – including basal RGCs, with both self-renewal and proliferative capacity, and more restricted progenitors. The stem cell-like properties of RGCs are linked to the fact that these cells are connected to the basal lamina, from which they receive proliferative signals via integrins. Now, Denise Stenzel et al. investigate in rodents the role of integrin αvβ3 in regulating the proliferative capacity of BPs ( Langerhans cells (LCs) are antigen-presenting cells of the epidermis, and play a key role in detecting pathogens in the skin and coordinating the immune response. However, the precursors of LCs during embryonic development are poorly characterised, particularly in humans. Here (
Langerhans cells (LCs) are antigen-presenting cells of the epidermis, and play a key role in detecting pathogens in the skin and coordinating the immune response. However, the precursors of LCs during embryonic development are poorly characterised, particularly in humans. Here (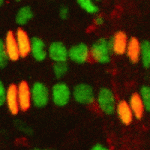 The Drosophila heart provides a relatively simple system for the analysis of gene regulatory networks (GRNs), as it comprises just two cell types – contractile cardial cells (CCs) and non-muscle pericardial cells (PCs). Moreover, many transcription factors (TFs) that regulate Drosophila heart development have been identified and shown to play conserved roles in mammals. On
The Drosophila heart provides a relatively simple system for the analysis of gene regulatory networks (GRNs), as it comprises just two cell types – contractile cardial cells (CCs) and non-muscle pericardial cells (PCs). Moreover, many transcription factors (TFs) that regulate Drosophila heart development have been identified and shown to play conserved roles in mammals. On  The shoot apical meristem (SAM) of higher plants contains the stem cell population that contributes to all above-ground organs. During vegetative growth, leaf primordia emerge from the periphery of the SAM, and the SAM transitions to an inflorescence meristem to initiate the reproductive phase. The WUSCHEL transcription factor specifies stem cell fate, and hence controls the size and activity of the SAM. A gene network involving the CLAVATA signalling pathway, the microRNA miR166g and HD-ZIPIII transcription factors is responsible for regulating WUSCHEL levels. On
The shoot apical meristem (SAM) of higher plants contains the stem cell population that contributes to all above-ground organs. During vegetative growth, leaf primordia emerge from the periphery of the SAM, and the SAM transitions to an inflorescence meristem to initiate the reproductive phase. The WUSCHEL transcription factor specifies stem cell fate, and hence controls the size and activity of the SAM. A gene network involving the CLAVATA signalling pathway, the microRNA miR166g and HD-ZIPIII transcription factors is responsible for regulating WUSCHEL levels. On 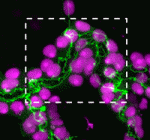 The Drosophila Mcr protein is a member of the thioester protein (TEP) family of proteins, which includes key regulators of innate immunity. Mcr is an unusual member of this family, possessing a putative transmembrane domain and lacking a key residue in the thioester motif. It has, nevertheless, been shown to be involved in phagocytic uptake in cultured Drosophila cells, although its putative immune functions have not been assessed in vivo. Two papers, from Stefan Luschnig and colleagues (
The Drosophila Mcr protein is a member of the thioester protein (TEP) family of proteins, which includes key regulators of innate immunity. Mcr is an unusual member of this family, possessing a putative transmembrane domain and lacking a key residue in the thioester motif. It has, nevertheless, been shown to be involved in phagocytic uptake in cultured Drosophila cells, although its putative immune functions have not been assessed in vivo. Two papers, from Stefan Luschnig and colleagues (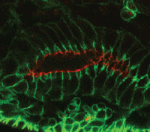 Both studies identify lethal EMS mutations inMcr in independent genetic screens, finding mutant phenotypes typical of SJ components. Consistent with a putative role in SJ formation, the protein colocalises with other SJ proteins at the lateral membrane. They further find that Mcr localisation is dependent on core SJ components and that, conversely, SJ proteins are mislocalised in Mcr mutants. At the morphological and ultrastructural level, SJs are disrupted in the absence of Mcr, and functional assays demonstrate that Mcr is required to form an effective paracellular barrier. As well as identifying a new SJ protein essential for barrier integrity, these two studies suggest an intriguing link between SJs and innate immunity. The epithelial barrier represents the first line of defence against pathogen invasion, andDrosophila haemocytes are known to undergo an epithelialisation-like process when encapsulating pathogens in the haemolymph. The identification of Mcr as a protein involved in both SJ formation and innate immunity now provides a molecular connection, and opens up new avenues for investigating potential functional links, between these two seemingly disparate processes.
Both studies identify lethal EMS mutations inMcr in independent genetic screens, finding mutant phenotypes typical of SJ components. Consistent with a putative role in SJ formation, the protein colocalises with other SJ proteins at the lateral membrane. They further find that Mcr localisation is dependent on core SJ components and that, conversely, SJ proteins are mislocalised in Mcr mutants. At the morphological and ultrastructural level, SJs are disrupted in the absence of Mcr, and functional assays demonstrate that Mcr is required to form an effective paracellular barrier. As well as identifying a new SJ protein essential for barrier integrity, these two studies suggest an intriguing link between SJs and innate immunity. The epithelial barrier represents the first line of defence against pathogen invasion, andDrosophila haemocytes are known to undergo an epithelialisation-like process when encapsulating pathogens in the haemolymph. The identification of Mcr as a protein involved in both SJ formation and innate immunity now provides a molecular connection, and opens up new avenues for investigating potential functional links, between these two seemingly disparate processes.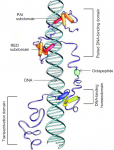 Pax genes encode a family of transcription factors that orchestrate complex processes of lineage determination in the developing embryo. Here, Judith Blake and Melanie Ziman review the molecular functions of Pax genes during development and detail the regulatory mechanisms by which they specify and maintain progenitor cells across various tissue lineages. See the Primer article on p.
Pax genes encode a family of transcription factors that orchestrate complex processes of lineage determination in the developing embryo. Here, Judith Blake and Melanie Ziman review the molecular functions of Pax genes during development and detail the regulatory mechanisms by which they specify and maintain progenitor cells across various tissue lineages. See the Primer article on p.  James Wells and Jason Spence review how recent advances in developmental and stem cell biology have made it possible to generate complex, three-dimensional, human intestinal tissues in vitro through directed differentiation of human pluripotent stem cells. See the Primer on p.
James Wells and Jason Spence review how recent advances in developmental and stem cell biology have made it possible to generate complex, three-dimensional, human intestinal tissues in vitro through directed differentiation of human pluripotent stem cells. See the Primer on p.  (No Ratings Yet)
(No Ratings Yet)