Perhaps, you believe it is important to make your expertise accessible to people—to scientists in other fields or those who are not in touch with scientific research at all. Or maybe you find your research work lonely or monotonous at times (that happens!) and you would like to do some fun stuff while engaging with others. Maybe, at the end of the day, your research funder obliges you to do some public outreach. And so, you decide you want to do some science communication. That’s great!
By the virtue of knowledge you have accumulated through years of studying and from first-hand experience of professional scientific research, you definitely have something to offer. The question is… Where do you start?
Very possibly, the phrase “science communication” immediately evokes particular associations for you. News outlets. Science fairs. Video blogs. Podcasts. Each one of these focusses on popularising science, discussing science or advocating for certain scientific topics. And so, it may seem then, that doing science communication means simply jumping on board one of those existing projects of your liking – or – starting your own such project by emulating one of them. And that would be a good start.
Still, let’s hang on for a moment. Take a breath. Think. What part of science would you like to talk about? This is not a trivial question. Science has so very many faces! Firstly, there are dozens of scientific disciplines and questions. Of course, there is your own research. However, oftentimes it is so narrow that it is impossible to talk about it without creating around it some comprehensible context. And that’s the first creative challenge to be mentioned.
Whether we want it or not, the specialised language – or jargon – we use in research is a product of a particular professional culture, the academic research culture, and jargon emerged to effectively operate in that culture. It would be naive to expect that someone outside the research world would be able to understand you without some pre-emptive induction or translation. Novice science communicators are often chided for excessive use of scientific slang and jargon. But really, the use of jargon is just a symptom of a bigger – and quite a fun and creative – challenge: how to bring closer and, ideally, organically blend the language of a particular scientific question with our everyday language?
The challenge becomes even more apparent as you widen your circle of discussion topics. Oftentimes, there is only so much you can say about one specific research problem, so, very likely, you would need to get comfortable talking about science that is not your own research: perhaps, something lateral to it or, maybe, different altogether. (Which is, again, normal since a narrow research topic very rarely satisfies the breadth of our own curiosity). And that’s great! Because that’s when you can clearly see that knowing something through research does not automatically translate into being able to make it understandable. What really helps is a certain attention to the creative possibilities of language, or, rather, languag-es we know and use, as well as our willfulness to explore those possibilities in practice. By languages, I don’t mean French, Cantonese or Swahili (although, it is helpful to remind ourselves that science is done in many languages and can – and should – be communicated in many languages too). I rather mean the different expressive and informational resources we use to communicate. One such example is the language of visuals. Or physical movement. Or – language of feelings, emotions and experiences.
Talking about emotions seems to be a sort of taboo in science. Still, that doesn’t mean that scientists don’t go through emotions or don’t experience things. For example, I think of motivation, surprise, wonder, happiness, frustration, boredom, doubt, disappointment, pessimism. On top of this, experiences are not erasable from research and research is not erasable from emotions, even if the (perceived) mark of the profession seems to be to distance oneself from them. That’s because, aside from being many other things, emotions are also our cognitive resources. They are not infallible – but neither is (mythical) “cold” reasoning – yet they help us grasp a way forward – or sideways – when there is no ready-made formula, method or plan, or when the existing ones don’t seem to work.
How does this all relate to the topic of science communication? Well, in the lab, field or library, we spend hours and hours chasing the phenomena we find curious or puzzling, going further and further (and further (still further)) down the rabbit hole of specifications, caveats, ruling-out contingencies and searching for parallels and convergences. This is quite the journey! The journey is full of uncertainties and surprises, which may or may not fully fade away eventually (e.g., think of the problem of induction). And as all this happens, at the very same time, people of other professions are engaged in their own journeys. Just like you may not have a clue about what they are up to, they too may not have a clue about the journey you and your colleagues go through in science. That’s precisely where another creative challenge lies. Sometimes, communicating science is about making it relatable, “experienceable”: understandable not merely as a commodified product, but as an activity, an experience, a journey.
To be clear, I’m not talking about a “hero’s journey”. Or about “constructing a compelling story.” As simple as it may sound, a story is propelled by experiences. But so is life. It is furnished with experiences of moving, staying, trying, avoiding trying, searching for and finding, or failing to find (huh?), or encountering the unexpected (wow!), not knowing what to do with, passing time, getting frustrated, forgetting (oh no!), connecting with, looking forward to and so on and so forth. We share experiences with each other as we find them entertaining, informative, useful, compelling, exciting, motivating, connecting, moving, and while sharing them, we call them “stories”. Navigating our way through first-hand or testimonial life experiences, we also use others’ stories to compose our own. Oftentimes we weave in metaphors or tropes to highlight this or that aspect. And, perhaps, this is how the gulf between the language of research and the language of everyday life can be traversed: via stories that sail back and forth and weave the two (three, five, seven) areas of experience together. To find – or to create – your way of doing this is a whole creative journey.
Some people may worry that storytelling can be dangerous: stories captivate, but they don’t contain an intrinsic filter for falsehoods. This is an enduring concern, but it is also somewhat undiscerning. On the one hand, it seems to imply that science – in practice or principle – is just about scientific facts and is devoid of imaginative leaps, tentative suggestions, discussions, unstructured reflections, detours and comebacks; always calm, clear, composed and certain. For all we know, this, in itself, is fictive, and a fiction not unproblematic. On the other hand, it is not clear what form of communication has intrinsic filters for falsehoods. Here one can start a long and tedious (or exciting) debate about forms of communication and metaphysics of truth, but the tentative answer I suggest is – none. Because, if we squint enough (enough, though), we may see stories as a form of technology; and, as with any piece of technology, it is the responsibility of the user – sensitive and attentive – to not mislead or deceive and, where necessary, to correct. Unfortunately, as Naomi Oreskes tells us, there are scientists who, from their position of authority, set forth rather harmful and deceptive stories. I imagine you, dear reader, are not interested in this path.
Still, it’s important to remember that science not only has many faces – but that those faces can look very different to different people. Perhaps, what you know as science – the insider’s view from the cockpit of your research domain – is your slice of science. You are surrounded by well-meaning researchers willing to positively contribute to society. However, when in 1972 the famous biophysicists Max Delbrück was asked whether pure science is to be seen as overall beneficial, he answered: “It depends”, and then added: “Clearly, the present state of the world – to which science has contributed much – leaves a great deal to be desired, and much to be feared”.
Today, this rings truer than ever. Partially because, pure or not, science heralds powerful technologies, and those, as we mentioned, do not always serve to the best ends. Which is why I personally like to remind myself that “science communication” is a shortening of a more accurate name: “public communication of science and technologies”. Science products – conceptual frameworks, technologies – often confront people where they are, sometimes unexpectedly, and it is not always a nice encounter. If this is what makes some people less trustful of some important scientific outputs, this distrust can hardly be remedied by filing more pamphlets with “correct” scientific answers. Which is why, the good practice of science communication often emphasises the importance of fostering connections, relations and dialogue.
And so we’re back at the creative challenges and the gulf between experiences and languages of science and everyday life. Only this time it is not just about the language and storytelling. It is also about meeting people where they are. Who are those people you want to connect with? What is your relation to them? Do you share the same concerns, worries or experiences? The same interests, cultural references or quirks? Same histories or aspirations? Where are they located? What is their preferred mode of communication? (And what if it’s not digital?) And how might they react to you, not just as a scientist, but as a fellow voyager through space and time and things and experiences? And as the traces of these questions evaporate into thin air, it is time to continue the halted action: to start exploring and experimenting with forms and formats of your science communication. Good luck!
Our November webinar featured two early-career researchers working on regeneration. Here, we share the talks from Stephanie Tsai (Massachusetts General Hospital) and Ben Cox (University of California, Davis).
We’ve launched a new preLights initiative: each month, preLighters with expertise across developmental and stem cell biology nominate a few recent developmental and stem cell biology (and related) preprints they’re excited about and explain in a single paragraph why. Short, snappy picks from working scientists — a quick way to spot fresh ideas, bold methods and papers worth reading in full. These preprints can all be found in the October preprint list.
Want to join us at preLights? If you’re keen to gain some science writing experience and be part of a friendly, diverse and international community, consider joining preLights and writing a preprint highlight article.
Have you ever wanted to image dozens of your favorite proteins, together, in 3D, and at single-cell resolution?
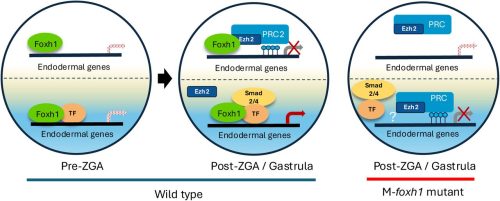
This preprint describes the development of a 3D adaptation of the previously described iterative indirect immunofluorescence imaging (4i) technique and its application to early zebrafish embryos to explore the heterogeneity in the onset of zygotic genome activation (ZGA). 3D-4i enables multiplexed immunofluorescence and in toto imaging of whole-mount structures, allowing high-resolution and high-throughput visualization of proteins and is supported by a comprehensive image analysis pipeline. Using 3D-4i, the authors capture the levels of proteins like cell cycle regulators, histone modifications, pluripotency factors, and RNA polymerase II at single-cell resolution. This leads to many key findings, including a framework to infer cell cycle phase and accurately predict transcriptional output, revealing how multiple features act collectively to precisely modulate ZGA onset.
A toolkit for testing membrane-localising tags across species Irene Karapidaki, Mette Handberg-Thorsager, Tsuyoshi Momose, Hitoyoshi Yasuo, Grigory Genikhovich, Sarah Assaf, Clara Deleau, Ying Pang, Clayton Pavlich, Beke Lohmann, Maria Lorenza Rusciano, Mattia Stranges, Juliette Mathieu, Marie Zilliox, Kirill Ustyantsev, Bastien Salmon, Béryl Laplace-Builhé, Manon Koenig, Jeffrey J. Colgren, Maria Ina Arnone, Eugene Berezikov, Thibaut Brunet, Gregor Bucher, Pawel Burkhardt, Daniel J. Dickinson, Evelyn Houliston, Jan Huisken, Lucas Leclère, Michalis Averof
preLight:
All things bright and beautiful.
Advances in technology have made detailed study of non-model organisms more feasible, yet there is always a challenge in applying existing techniques to new systems. The authors take a systemic approach to screening a varied set of membrane-localisation tags in the early embryos of a wide range of organisms. They identify several tags that display strong, membrane specific fluorophore localization in many species but highlight that no single tag is ubiquitously successful. This work is an open science project that combines the efforts of many labs to provide a useful community resource. Check this preprint out if you’re looking to label membranes in your species of interest!
Post-translational Tuning of Human Cortical Progenitor Neuronal Output Julien Pigeon, Tamina Dietl, Myriame Abou Mrad, Ludovico Rizzuti, Miguel V. Silva, Natasha Danda, Corentine Marie, Clarisse Brunet Avalos, Hayat Mokrani, Laila El Khattabi, Alexandre D. Baffet, Diogo S. Castro, Carlos Parras, Boyan Bonev, Bassem A. Hassan
preLight:
A novel role for PTMs in fine tuning neurogenesis.
While human brain development has long been linked to alterations in genomic sequence, the authors raise the question: do post translational modifications (PTMs) offer a complementary mechanism in shaping human brain evolution ? In particular, they focus on Neurogenin 2 (NEUROG2), a master regulator of neural fate and neuronal identity specification, whose activity is dependent on PTMs such as phosphorylation. Through a combination of genome editing, high-throughput imaging, and single-cell multiomics, they investigate whether NEUROG2 has evolved species-specific functional plasticity in human radial glial cells (RGCs). They find that the human NEUROG2 regulates both deep and upper layer neuron production and controls the balance between proliferative and neurogenic divisions in RGCs via its phosphorylation at residue T149. This phosphorylation tunes AP-1 (JUN/FOS) driven gene regulatory networks in RGCs, enhancing neurogenic commitment and increasing upper-layer neuron production. Phospho-mutant NEUROG2 promotes premature chromatin opening at AP-1 binding sites, priming RGCs for differentiation without accelerating neuron maturation. Overall, their findings suggest that the evolutionary innovations in brain development do not solely rely on genetic changes but can also arise through modifications of conserved proteins.
Shark and Salamander – pioneers in building beautiful brains.
“Evolutionary change is often driven by changes in development.”
Most of our understanding of brain development comes from studying the mouse (mammal). However, a tubular brain and backbone are the defining features of a subphylum – vertebrata. What if mammalian brain development actually combines elements found in the brain developmental programs of fish, salamanders, and birds, much like a musical genre blending influences from several styles? These two papers dig into shark and salamander brains using a combination of single-cell RNA sequencing, spatial transcriptomics, birth-dating, lineage tracing, and computational approaches to learn the origin of brain development as we know it.
Combining these two papers is as satisfying as putting together a jigsaw puzzle.
The shark brain has field-level homology with that of the salamander and mouse.
Both sharks and salamanders have multipotent progenitors that give rise to intermediate progenitors: the driving force behind big brains.
The Cajal-Retzius cells marked their enigmatic presence in sharks.
In salamander, the molecular identity, layer position, and projection are functions of birthdate.
With this prologue, dive into the many observations these two papers make, and discuss where the point of difference arose in the vertebrates that made their brains look and behave differently.
IPSC-based modeling of resiliency in centenarians reveals longevity-specific signatures Todd W. Dowrey, Samuel F. Cranston, Nicholas Skvir, Yvonne Lok, Payton Bock, Elizabeth K. Kharitonova, Elise MacDonald, Ella Zeldich, Christopher Gabel, Alexander Tyshkovskiy, Stefano Monti, Vadim N. Gladyshev, Paola Sebastiani, Thomas T. Perls, Stacy L. Andersen, George J. Murphy
preLight:
Giving new life to elderly cells reveals what makes them resilient to aging.
Understanding the molecular and cellular mechanisms that govern aging has been, and will likely remain, a central question for humanity. What better way to approach this challenge than by studying individuals who appear to defy canonical aging mechanisms? In this study, the authors generated pluripotent stem cells from a cohort of centenarians and differentiated them into excitatory cortical neurons. Molecular and cellular comparisons with neurons from non-centenarian individuals revealed that centenarian-derived neurons exhibit a distinct resilience signature, marked by enhanced synaptic integrity, calcium homeostasis, and energy-efficient metabolism at baseline. When challenged, these neurons demonstrated superior dynamic stress responses, in contrast to non-centenarian neurons, which showed chronic proteostatic stress activation and blunted responsiveness. Overall, this work highlights the versatility of the stem cell platform in uncovering molecular mechanisms that confer resilience to aging in neural systems. This represents a foundational resource for investigating the determinants of aging across diverse cell types and developmental contexts, leveraging the innate ability of stem cells to recapitulate key human developmental processes.
Decoding mammalian body axis elongation: a supracellular ‘actin cap’ in action.
Body axis elongation is fundamental to establishing a head-to-tail body plan in vertebrates, including mammals. Although the genetic and biochemical pathways involved are well studied, the physical forces that help shape the mammalian axis remain less understood. To investigate these mechanisms, the authors of this preprint used mouse and human stem-cell-derived gastruloids, an accessible model that bypasses the challenges of working with embryos in utero. By integrating a previously developed gastruloid analysis framework and oil droplet-based deformation measurements, the authors aimed to uncover the mechanical forces at play. Their findings show that randomly oriented cell divisions generate isotropic expansive forces throughout the gastruloid during the elongation period. However, a posteriorly enriched actin network, termed the ‘actin cap’, provides localized mechanical resistance, preventing tissues at the posterior domain from expanding laterally, thereby guiding the elongation of the body axis. Apart from mouse and human gastruloids, mouse embryo explants display similar proliferation and actin patterns, supporting the idea that this actin cap–based mechanical constraint is a conserved and previously overlooked mechanism in mammalian axis elongation.
What if we could use patient cells to generate neurons that can replace dysfunctional native cells and tackle diseases characterised by the aberration of atrial electromechanical activity, namely atrial fibrillation?This is the question the authors of this manuscript in preprint answer, a detailed protocol for the derivation of parasympathetic neurons from human induced pluripotent stem cells (hiPSC). The described protocol includes several useful features, including the lack of batch-testing for growth factors as well as the integration of electrophysiological and functional assessment testing to specifically identify the presence of parasympathetic neurons. The authors also describe cellular features the user should look out for, to ensure proper progression through the protocol steps, including the presence of smooth spheroids during the Embryoid bodies stage and the presence of neuronal-like projections during the neuronal differentiation stage, the expression of specified markers of autonomic (ASCL1, PHOX2B) and parasympathetic (CHAT, VACHT) populations and markers of autonomic neuron development (ISL1). The protocol also includes troubleshooting sections, which is sure to help new users make the most of it.
Water must flow, but will blood in in vitro models do the same? In this study, the authors generate endothelial cells (EC) derived from human induced pluripotent stem cells (hiPSC); efficiency of hiPSC-derived EC generation is enhanced via overexpression of ETV2, a factor involved in vascular and cardiac development. In short, expression of ETV2 is induced in the hiPSC lines used; these are then subjected to differentiation protocols that will eventually generate EC. In vitro, these same cell lines can self-assemble into stable and lumenized microvascular networks (MVN) on the surface of microfludic chips; more importantly, however, no such success in vascular formation has been observed in lines subjected to the conventional differentiation models, highlighting the importance of growth factor overexpression in pluripotent source populations. This study provides an answer to the problem of organoid vascularization and can be applied in models examining tumor vascularization as well as models evaluating the blood brain barrier (BBB).
Spotted a preprint in this list that you love? If you’re keen to gain some science writing experience and be part of a friendly, diverse and international community, consider joining preLights and writing a preprint highlight article.
Note: A group of preLighters, with expertise across developmental and stem cell biology, have highlighted (in orange) their favourite preprints of this month. Check out the accompanying post to learn why they picked these articles.
Post-translational Tuning of Human Cortical Progenitor Neuronal Output Julien Pigeon, Tamina Dietl, Myriame Abou Mrad, Ludovico Rizzuti, Miguel V. Silva, Natasha Danda, Corentine Marie, Clarisse Brunet Avalos, Hayat Mokrani, Laila El Khattabi, Alexandre D. Baffet, Diogo S. Castro, Carlos Parras, Boyan Bonev, Bassem A. Hassan
A human arteriovenous differentiation roadmap reveals vein developmental mechanisms and vascular effects of viruses Lay Teng Ang, Sherry Li Zheng, Kevin J. Liu, Anastasiia Masaltseva, June Winters, Isabel von Creytz, Sawan K. Jha, Qingqing Yin, Crystal Qian, Xiaochen Xiong, Amir Dailamy, Ellie Xi, Juan C. Alcocer, Daniel W. Sorensen, Richard She, Karina Smolyar, Dorota Szumska, Svanhild Nornes, Renata M. Martin, Benjamin J. Lesch, Nicole K. Restrepo, Wenfei Sun, Jonathan S. Weissman, Heiko Lickert, Matthew P. Porteus, Mark A. Skylar-Scott, Christian Mosimann, Saulius Sumanas, Sarah De Val, Joseph B. Prescott, Kristy Red-Horse, Kyle M. Loh
Induction of menstruation in mice reveals the regulation of menstrual shedding Çağrı Çevrim, Nicholas J. Hilgert, Aellah M. Kaage, Andrew J.C. Russell, Allison E. Goldstein, Claire J. Ang, Jaina L.R. Gable, Laura E. Bagamery, Ana Breznik, Daniela J. Di Bella, Mustafa Talay, Jingyu Peng, Kathleen E. O’Neill, Fei Chen, Sean R. Eddy, Kara L. McKinley
Domain-specific mechanisms of YAP1 variants in ocular coloboma revealed by in-vitro and organoid studies Srishti Silvano, Annika Rick-Lenze, James Bagnall, Mrinalini Saravanakumar, Xinyu Yang, Robert Lea, Lindsay Birchall, Julie R. Jones, Jessica M. Davis, Anzy Miller, Rachel E. Jennings, Elliot Stolerman, Jamie M. Ellingford, Simon C. Lovell, Forbes Manson, Gavin Arno, Panagiotis I. Sergouniotis, Cerys S. Manning
IPSC-based modeling of resiliency in centenarians reveals longevity-specific signatures Todd W. Dowrey, Samuel F. Cranston, Nicholas Skvir, Yvonne Lok, Payton Bock, Elizabeth K. Kharitonova, Elise MacDonald, Ella Zeldich, Christopher Gabel, Alexander Tyshkovskiy, Stefano Monti, Vadim N. Gladyshev, Paola Sebastiani, Thomas T. Perls, Stacy L. Andersen, George J. Murphy
Maternal transmission of a plastid structure enhances offspring fitness Tyler J. Carrier, Andrés Rufino-Navarro, Thorben Knoop, Urska Repnik, Andrés Mauricio Caraballo-Rodríguez, David M. Needham, Corinna Bang, Sören Franzenburg, Marc Bramkamp, Willi Rath, Arne Biastoch, José Carlos Hernández, Ute Hentschel
A toolkit for testing membrane-localising tags across species Irene Karapidaki, Mette Handberg-Thorsager, Tsuyoshi Momose, Hitoyoshi Yasuo, Grigory Genikhovich, Sarah Assaf, Clara Deleau, Ying Pang, Clayton Pavlich, Beke Lohmann, Maria Lorenza Rusciano, Mattia Stranges, Juliette Mathieu, Marie Zilliox, Kirill Ustyantsev, Bastien Salmon, Béryl Laplace-Builhé, Manon Koenig, Jeffrey J. Colgren, Maria Ina Arnone, Eugene Berezikov, Thibaut Brunet, Gregor Bucher, Pawel Burkhardt, Daniel J. Dickinson, Evelyn Houliston, Jan Huisken, Lucas Leclère, Michalis Averof
Systematic Review of Over A Century of Global Bioscience Research Okechukwu Kalu Iroha, Dauda Wadzani Palnam, Peter Abraham, Israel Ogwuche Ogra, Ndukwe K. Johnson, Elkanah Glen, Dasoem Naanswan Joseph, Seun Cecilia Joshua, Grace Peter Wabba, Morumda Daji, Dogara Elisha Tumba, Mercy Nathaniel, Emohchonne Utos Jonathan, Samson Usman, Mela Ilu Luka, Vaibhav B. Sabale, Emmanuel Oluwadare Balogun, Umezuruike Linus Opara
I was so excited when I received notification that my first first-author research paper was accepted. My excitement quickly turned into sadness with the realization that my co-PI was not seeing our vision come to fruition.
The first time I met Scott Hawley was in my genetics module. As a fresh PhD student at the Stowers Institute for Medical Research, I was eager to soak up new knowledge. Studiously, I had already gone through the recommended reading material, chapters from one of Scott’s books. Post it notes were poking out of my binder containing those pages, revealing where I had written down thoughts and questions. The idea of Scott could be intimidating: an infamous scientist in the fields of meiosis and fly genetics. Quickly it became clear that there was no reason to be intimidated. He had a passion for teaching and supporting others where he could. Shortly after we wrapped up the genetics module, I ran into Scott in the cafeteria where he asked if I would consider rotating in his lab. I was exhilarated because I already had the same idea.
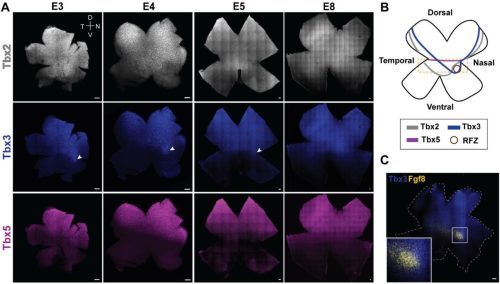
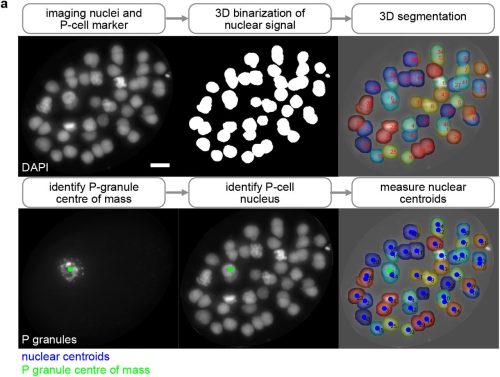
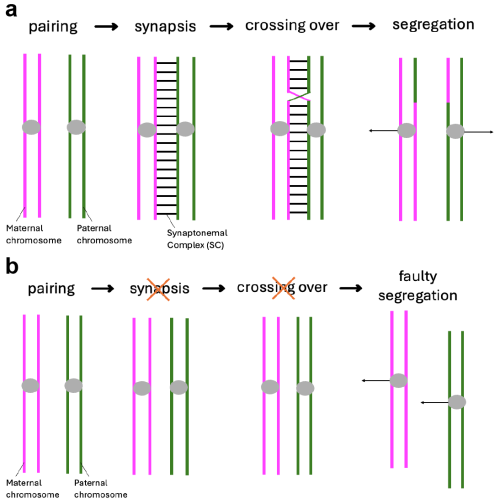
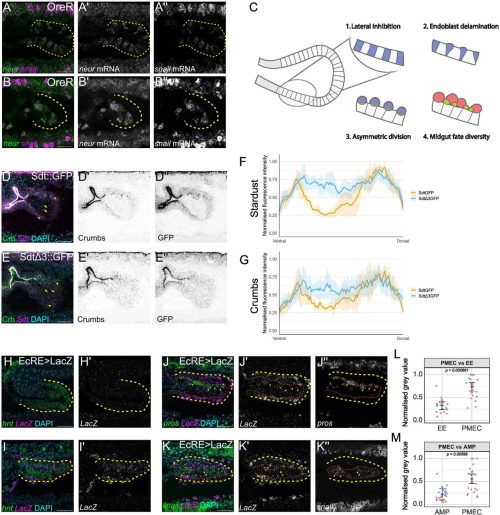
When I started my first lab rotation a few months later, I was brimming with what we in Germany would call “Vorfreude” (engl.: “pre-joy”). I had previously met with Scott and my supervisor Cathy about my project for the next eight weeks. The project they had in mind for me was focused on the synaptonemal complex, a structure I had never heard of before coming to Stowers. The topic sounded exciting and I quickly became fascinated by it. What I came to understand was that in early meiosis, this protein structure forms between the paired maternal and paternal copy of each chromosome (Fig. 1a). If it does not form properly, crossovers between these two chromosome copies cannot be made properly, causing the faulty segregation of chromosomes (Fig. 1b). The lab of Scott Hawley was one of the many labs trying to understand how the synaptonemal complex was involved in these processes.
Fig. 1: Meiosis I and the synaptonemal complex (SC). a) Under normal conditions, the maternal and paternal copy of each chromosome pair and the SC subsequently forms between them. This allows for the formation of crossovers between the homologous chromosomes which ensures their proper segregation at the end of meiosis I. b) If the SC does not form properly, crossovers are severely diminished causing faulty chromosome segregations.
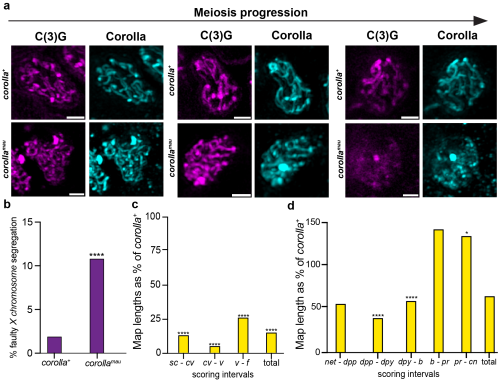
The Hawley lab, and one of my co-authors specifically, had established a mutated fruit fly. This was nothing out of the ordinary for fly people, we love that stuff! This interesting mutation removed the gene coding for the synaptonemal complex protein Corolla and inserted in that very location the coding sequence for the same protein but from a closely related species. My goal for the rotation was to perform the first characterizations of the effects this gene replacement had on meiosis. Cathy already had a plan for which experiments I should do and with her support I got to work. I made crosses, scored flies, and performed microscopy. Some of the work was old habit, most of it was new to me. Quickly it became clear what we were dealing with: The synaptonemal complex in these flies was disassembling too early (Fig. 2a) and, unsurprisingly, there was a moderate but significant increase in faulty chromosome segregation (Fig. 2b)1. Where it got confusing (and intriguing!) was that crossover rates on the X chromosome were almost entirely abolished (Fig. 2c)1. The near complete lack of X chromosome crossovers should lead to a dramatically higher rate of errors in chromosome segregation. Scott loved it and he offered me to join his lab to figure out how these phenotypes arose.
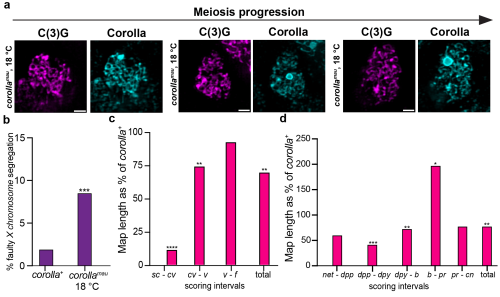
Fig. 2: The effect of the corolla gene replacement on meiosis. a) Microscopy images of the SC in wild type fruit flies (= corolla+)and the gene replacement flies (= corollamau) visualized through immunofluorescence of the two SC proteins C(3)G and Corolla. While the SC stays intact through meiotic progression in corolla+ flies, it disassembles in corollamau flies. Scale bar represents 1 mm length. b) Rate of faulty X chromosome segregation. corollamau flies have an increased rate of chromosome segregation errors. c) Crossovers on the X chromosome as a percentage of rates in corolla+ flies. Crossover rates are significantly lower. d) Crossovers on the 2ndchromosome as a percentage of rates in corolla+ flies. Specific intervals show a decrease or increase of crossovers but overall there is no significant difference. * means p ≤ 0.05, ** means p ≤ 0.01, *** means p ≤ 0.001, **** means p ≤ 0.0001.
A bit less than 16 weeks later I had made my decision. While I was not going to join the Hawley lab as he first imagined it, I was going to be co-supervised by Scott Hawley and Matt Gibson to work on a joint project. However, we agreed that I could continue working on the project I had begun during my rotation in the Hawley lab. The experiments should have been straightforward but as science often goes, it took longer than we thought to finish. As a clear next step, I looked at the rates of crossovers on a different chromosome. Fruit flies only have four chromosomes: The sex chromosomes X and Y, the two large autosomes as known as the 2ndand 3rdchromosomes, and a very small 4th chromosome. We needed to look at one of the large autosomes. It would not have made a huge difference as we knew there was differences in how crossovers on the sex chromosomes and autosomes respond to synaptonemal complex defects but autosomes between one another did not show major differences2. Therefore, we decided to look investigate crossovers on the 2ndchromosome as a representative autosome. While crossovers were almost entirely absent from the X chromosome, the effect on crossovers on the 2ndchromosome was nowhere near as dramatic (Fig. 2d)1. These results solved our confusion as to why the chromosome segregation machinery was not overwhelmed by chromosomes lacking crossovers.
Thinking the work was done, I presented the story at an internal seminar. As it was my first talk in front of a larger audience, it was exhilarating, especially because I received a lot of questions at the end of my talk. One of those questions was fascinating: “Have you tried looking at these flies at a colder temperature? That might change its disassembly dynamics.” Scott loved the idea and it was an easy experiment to do. The standard rearing temperature we maintained the flies at was 25 °C. I moved these flies to 18 °C instead and indeed, the synaptonemal complex stayed fully intact (Fig. 3a)1! So, I went back to the bench to repeat the chromosome segregation and crossover experiments at this colder temperature. Due to the rescue of the early disassembly of the synaptonemal complex, crossovers on the X chromosome were improved but overall crossovers still did not reach wild type levels and chromosome segregation was still significantly higher than in wild type flies (Fig. 3b-d)1. This was even more exciting as it meant that this synaptonemal complex protein, Corolla, is not only part of the structure itself but directly involved in crossover formation.
Fig. 3: Temperature-sensitive phenotypes in corollamau flies. a) Microscopy images of the SC in wild type fruit flies (= corolla+)and the gene replacement flies (= corollamau) visualized through immunofluorescence of the two SC proteins C(3)G and Corolla. The SC stays intact throughout meiotic progression in corollamau flies reared at 18 °C. Scale bar represents 2 mm length. b) Rate of faulty X chromosome segregation. corollamau flies reared at 18 °C still have an increased rate of chromosome segregation errors. c) Crossovers at 18 °C on the X chromosome as a percentage of rates in corolla+ flies. Crossover rates are significantly lower. d) Crossovers at 18 °C on the 2ndchromosome as a percentage of rates in corolla+ flies. Specific intervals show a decrease or increase of crossovers but overall there is no significant difference. * means p ≤ 0.05, ** means p ≤ 0.01, *** means p ≤ 0.001, **** means p ≤ 0.0001.
I was finishing up some last experiments before writing up the complete story. During that time, on a Friday morning in late January I arrived at the lab and Cathy quickly found me. By the look of her face, I immediately knew that she was bearing bad news. Scott had died that morning. Of course, in good old Scott fashion, he had emailed people from the lab about work related matters earlier that morning just prior to that. I liked that. I think that’s how he would have wanted to go, thinking of his science until the very end. Him being gone did not make any sense. I couldn’t even imagine how Cathy must have felt. She has worked with Scott for over two decades. Even for me, Scott was more than just my co-PI. He was my biggest supporter, always looking for ways to push me and my career further. We shared ideas for crazy experiments and talked about papers we recently read. I had realized already that I wanted to continue studying the synaptonemal complex beyond my PhD, a field I stumbled into by accident. And while the fact that he to his last day suffered from imposter syndrome did not fill me with confidence that I could beat mine, a part of me felt like I could make it because he said so. But without him? I was not so sure anymore.
I don’t remember much of what I did the following couple of months, but I did finish those last experiments. With the support of Cathy and Stacie, another long-term Hawley lab scientist, I was able to write up the story for the first time. Looking back, I will always remember the embarrassing first draft I sent them. Cutting myself some slack, it was my first time writing a scientific paper. What I came to appreciate is that a first draft is better than nothing because editing is a lot easier than writing about something for the first time. After several drafts went through multiple people (thank you all!), the manuscript was done. The first submission was desk-rejected, which was frustrating, but the second attempt went better. One round of very fair but major revisions later and the paper was accepted.
The acceptance of a scientist’s first first-author publication in a peer-reviewed journal is a milestone achievement. Regardless of what your paper is about, you had to overcome a multitude of hurdles to get to this point. When I saw the notification, I was filled with so much joy. I shared the news with my co-authors, colleagues, and friends. Everyone was excited! Then the sadness hit me. The one person who will not be celebrating this achievement with me was Scott. At that point it had been almost exactly nine months since he passed. I had accepted his death though certain situations were still upsetting. For some reason I hadn’t considered that this particular moment would upset me as well. It was his exuberant joy which I was able to channel and get out of this. I could imagine the email he would send in response to the manuscript acceptance and his palpable jubilation when we would have celebrated in person. I always knew that it was not me who was special but that he made everyone feel special in those moments, something that I can only imagine stems from his own imposter syndrome. There are plenty of people who will celebrate the very big papers and talk admiringly about their favorites. Scott didn’t care where your paper ended up and every trainee who went through his lab was “one of his best”. It is exactly that skill of Scott’s to make everyone around him feel important and special, that has made it possible for me to accept those bittersweet moments. If I can carry only a fraction of that faith forward, into how I move through my own work, how I teach, and how I mentor, then some part of Scott’s spirit will continue living through me and hopefully inspire others.
Every human face is unique, allowing us to distinguish between individuals. We know little about how facial features are encoded in our DNA, but we may be able to learn more about how our faces develop by looking at our ancient relatives, the Neanderthals. Neanderthal faces were quite distinctive from our own, with large noses, pronounced brows and a robust lower jaw. Now, scientists from the MRC Human Genetics Unit in the Institute of Genetics and Cancer at the University of Edinburgh, UK, are using the DNA of our extinct distant relatives to learn more about how faces develop and evolve. Published today in the journal Development, they show how a region of Neanderthal DNA is better at activating a jaw-forming gene than the human counterpart, revealing one potential reason for Neanderthal’s larger lower jaws.
Hannah Long (University of Edinburgh, UK), who led the study, explains that scientists have sequenced the Neanderthal genome using DNA extracted from ancient bone and says, “The Neanderthal genome is 99.7% identical to the genome of modern-day humans and the differences between species are likely responsible for altering appearance”. Both human and Neanderthal genomes consist of about 3 billion letters that code for proteins and regulate how genes are used in the cell, which makes finding regions that impact appearance like looking for a needle in a haystack. Fortunately, Long and her colleagues had an informed idea where to look first: a region of the genome that is linked to Pierre Robin sequence, a syndrome in which the lower jaw is disproportionately small. “Some individuals with Pierre Robin sequence have large deletions or DNA rearrangements in this part of the genome that change face development and limit jaw formation. We predicted that smaller differences in the DNA might have more subtle effects on face shape,” said Long.
By comparing human and Neanderthal genomes, the team found that in this region, roughly 3000 letters in length, there were just three single-letter differences between the species. Although this region of DNA doesn’t contain any genes, it regulates how and when a gene is activated, specifically a gene called SOX9, a key coordinator of the process of face development. To demonstrate that these Neanderthal-specific differences are important for the development of the face, Long and colleagues needed to show that the Neanderthal region could activate genes in the right cells at the right time as the embryo develops. The researchers simultaneously inserted the Neanderthal and human versions of the region into the DNA of zebrafish and programmed the zebrafish cells to produce different colours of fluorescent protein depending on whether the human or Neanderthal region was active. Watching the zebrafish embryos develop, the researchers found that both the human and Neanderthal regions were active in the zebrafish cells that are involved in forming the lower jaw and the Neanderthal region was more active than the human version.
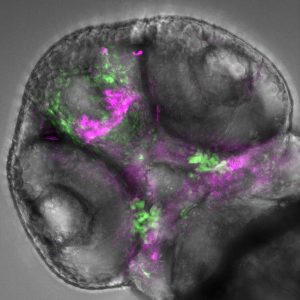
A 2-day-old zebrafish embryo viewed from below; with the zebrafish head pointing towards the top-left corner of the image. Within the transparent embryo, the fluorescent cells show activity of the inserted human DNA in specific cells (lower green signal) overlapping with the developing jaw (lower purple signal). Image credit: Kirsty Uttley and Hannah Jüllig.
“It was very exciting when we first observed activity in the developing zebrafish face in a specific cell population close to the developing jaw, and even more so when we observed that the Neanderthal-specific differences could change its activity in development,” said Long. “This led us to think about what the consequences of these differences could be, and how to explore these experimentally.” Knowing that the Neanderthal sequence was more powerful at activating genes, Long and colleagues then asked if the resulting increased activity of its target, SOX9, might change the shape and function of the adult jaw. To test this theory, they provided the zebrafish embryos with extra SOX9 and found that cells that contribute to forming the jaw occupied a larger area.
“In our lab, we are interested in exploring the impact of additional DNA sequence differences, using a technique that mimics aspects of facial development in a dish. We hope this will inform our understanding of sequence changes in people with facial conditions and inform diagnosis,” says Long. This research shows that by studying extinct species we can learn how our own DNA contributes to face variation, development and evolution.
Uttley, K., Jüllig, H. J., De Angelis, C., Auer, J. M. T., Ozga, E., Bengani, H. and Long, H. K. (2025). Neanderthal-derived variants increase SOX9 enhancer activity in craniofacial progenitors that shape jaw development. Development, 152, dev204779. doi:10.1242/dev.204779
Last September marked the return of our image competition with the MBL Embryology course at Woods Hole. The 2025 cohort submitted impressive images, ranging from polychaete worms to butterflies, squid, and mice, using a range of microscopy techniques. Here, we interview Nicole Roos and Anthony Wokasch, the winners of the popular vote of the image competition with their submission, ‘Mouse embryo’. As winners of the image competition, their submission was featured on the cover of a recent issue of Development.
Can you describe your research career so far?
Nicole: As a kid I loved conducting at-home experiments, visiting the science museum and attending my chemistry and biology classes, so pursuing a career in science has always been a no brainer for me. I started my research career as a freshman at The University of Texas at Dallas, USA, while pursuing my BSc in Biochemistry. Inspired by a family member who survived breast cancer, I joined the lab of Dr Nikki Delk to study chronic inflammation in tumor microenvironments. My interest in genetics was sparked while investigating mutations that affect chromatin remodeler function at the University of Texas Southwestern Medical Center, USA, with Dr Laura Banaszynski. Now, I am a fourth year PhD candidate working with Dr Leila Rieder in the Genetics and Molecular Biology program at Emory University, USA.
Anthony: I began my research career at American University, USA, where I completed a joint BSc/MSc in Biology. During my time there, I did my Master’s thesis in the lab of Naden Krogan, where I characterized the role of the floral boundary gene SUPERMAN (SUP) in a transcriptional mechanism which regulates gene expression and floral organ patterning of the reproductive organs in the flower of Arabidopsis thaliana. Following completion of my Master’s, I became a fellow at the National Cancer Institute (NCI) in the lab of Dr Peter D. Aplan. There I tested the effects of a DNA methyltransferase (DNMT1) inhibitor in a murine model for Myelodysplastic Syndrome (MDS). However, at this point my interest in development was overwhelming and led me to pursue a PhD at Vanderbilt University, USA, where I am a PhD candidate in my fourth year.
Can you tell us about your current research?
Nicole: At Emory, I use the powerful model system Drosophila melanogaster (fruit flies) to study how histone genes, which encode proteins that package and organize DNA in the nucleus, are regulated in embryogenesis and throughout development. Histone gene regulation is uniquely regulated throughout development, and this regulation is carried out by a nuclear body called the histone locus body. Not only do I get to study genetics, but through my research I took interest in developmental biology. This compelled me to apply to the Embryology course offered by the Marine Biological Laboratory (MBL) in Woods Hole, USA, where I discovered a strong desire to study regeneration and/or evolutionary developmental biology in the future.
Anthony: My current research, under the guidance of Dr Maureen Gannon focuses on how the Pdx1 transcription factor and its C-terminal interacting factors, Oc1 and SPOP, regulate the choice between proliferation or differentiation of early pancreatic progenitor cells. This work has deepened my understanding of how epigenetic and molecular factors control cell fate decisions during organogenesis.
What is your favourite imaging technique/microscope?
Nicole: The majority of my research at Emory University requires widefield fluorescence microscopy, so I was excited to take the Embryology Course at the MBL to learn various confocal microscopy techniques. I quickly became fascinated with live imaging using various spinning disc confocal microscopes (my favorite being the Nikon Yokogawa W1 spinning disc) and collaborated with other students to live image processes such as sea urchin egg and sea star sperm cross-fertilization (sea sturchins), nematode egg laying, and early embryonic cleavage of comb jellies. However, I also loved taking intricate fluorescence images on the Evident FV400 scanning confocal, the same microscope we used to take the image of the stained mouse embryo.
Anthony: My favorite microscope is the Evident FV4000 Confocal Laser Scanning Microscope, which we used at the MBL during the course to take this is image.
What are you most excited about in microscopy?
Nicole: I’m fascinated by current microscopy techniques which bypass diffraction limit imaging and visualize single molecule interactions. The development of expansion microscopy techniques alongside super resolution microscopy allow scientists to visualize cell biology more deeply now than ever before. I am especially intrigued by the combination of expansion microscopy with stimulated electron depletion (STED), which has been used in fluorescence imaging of chromatin at the nucleosomal level. This can inform the field of developmental biology and my field, where we study DNA-protein interactions to understand how histones are regulated.
Anthony: I am really excited right now doing whole-mount immunofluorescence of embryonic mouse pancreas samples and using confocal microscopy and or light-sheet (LSFM) to reveal key changes of cell fate choices and morphology during development.
Our October webinar featured two early-career researchers working on development across scales. Here, we share the talks from Osvaldo Contreras (Victor Chang Cardiac Research Institute and UNSW) and Yinan Wan (Biozentrum, University of Basel).
[Editorial from Development’s latest Special Issue – Lifelong Development: the Maintenance, Regeneration and Plasticity of Tissues, edited by Merixtell Huch and Mansi Srivastava.]
Traditionally, developmental biology has been considered the study of the embryo, and significant events such as metamorphosis or birth signify the pinnacle of development. However, we now better appreciate that development is a continuum and that – as in plants – developmental processes occur throughout the lifetime of an animal. Cell fate specification and differentiation, morphogenesis and patterning can continue after embryonic development; growth, degrowth, ageing, regeneration, and even reverse development (e.g. in some disease states) are just some examples of development-like processes occurring during the life history of a species. This special issue sought to draw these connections and to highlight how embryonic studies have revealed fundamental lifelong principles, advocating for a broader interpretation of developmental biology that circumvents restriction to specific stages in the life cycle.
The 26 research and review-type papers published in this issue illustrate this goal, including a breadth of mechanisms, research organisms and organ types. A selection of Research Articles demonstrates how several tissues and organs continue to develop, differentiate and mature during post-embryonic stages, establishing the principle of lifelong development. These examples include the mouse gut (Pan et al., 2025) and adipose tissue (Mahapatra et al., 2025), zebrafish vasculature (Preußner et al., 2025), and the Caenorhabditis elegans germline (Gupta et al., 2025). The mammalian nervous system represents a particularly well-studied example of postnatal refinement in response to sensory stimuli and learning. Therefore, we are glad to capture studies that discuss the ongoing development of retinal cells (Shah et al., 2025) and visual cortex (Xavier et al., 2025), microglia (Hammond et al., 2025), astrocytes (Iyer et al., 2025) and neurons (Liu et al., 2025). In addition to an extension of embryonic development, some tissues and organs undergo extensive remodelling during metamorphosis in invertebrates or puberty in humans (Rauner et al., 2025), and tissue-resident stem cells are crucial for the homeostasis and maintenance of adult tissues, which may also change behaviour over a lifespan due to shifting niche environments (Puri and Blanc, 2025).
Beyond these examples of post-embryonic development, regeneration offers perhaps the most striking illustration of development-like processes occurring in adults. We are therefore excited that regenerative studies are well represented in the special issue. Research papers characterise the initial molecular events and signalling in regeneration (Quinn et al., 2025), as well as the regeneration of specific tissues and organs, such as the zebrafish skin (Craig et al., 2025) and heart (Feng et al., 2025; Forman-Rubinsky et al., 2025). Whole-body regeneration is also explored in planarians, with studies revealing how these species maintain robust regenerative potential throughout life (Zelko et al., 2025), and the mechanisms by which polarity and patterning are re-established during regeneration (Anderson and Petersen, 2025; Miliard et al., 2025). In addition, our review-type content highlights a recent workshop from this field (Bayin et al., 2025), the interplay between vertebrate regeneration and the nervous system (Wakelin and Johnston, 2025; Tendolkar and Mokalled, 2025) and non-traditional model systems with remarkable regenerative potential (García-Arrarás et al., 2025). We are pleased that one of these organisms, the tapeworm, also features in a Research Article in the same issue (Nanista et al., 2025). Moreover, our Techniques and Resources articles provide valuable references for studying adult stages of highly plastic species (Temiz et al., 2025; Little et al., 2025), with a Hypothesis article exploring such phenotypic plasticity as the basis of complex developmental potential (Dardiry and Ikmi, 2025).
This special issue underscores the continuity of developmental processes across the lifespan from embryogenesis to regeneration, tissue maintenance and phenotypic plasticity. By linking classical developmental biology and emerging insights into post-embryonic and adult stages, we aim to broaden the field’s conceptual understanding of developmental biology. We hope you enjoy reading the issue and that it inspires further investigation of how developmental mechanisms operate beyond early life, adapt to environmental cues, and contribute to lifelong organismal plasticity. Development welcomes future submissions that explore these dynamic and evolving aspects of development across diverse systems and life stages.
Spotted a preprint in this list that you love? If you’re keen to gain some science writing experience and be part of a friendly, diverse and international community, consider joining preLights and writing a preprint highlight article.
Yuan-Chen Tsai, Hajime Ozaki, Xinyi Wang, Axel A. Almet, Isabel Fleming, Kaori Shiraiwa, Matthew Jung Min Noh, Caihao Nie, Sunnyana Trejo, Bret Kiyoshi Sugita, Jiya Dalal, Ruben Alberto Gonzalez, Briana De Jesus, Gregory Li-Min Chen, Michael J Gandal, Qing Nie, Momoko Watanabe
Wilke H. M. Meijer, Virginia Andrade, Suzan Stelloo, Wouter M. Thomas, Marek J. van Oostrom, Eveline F. Ilcken, Kim T. J. Peters, Michiel Vermeulen, Katharina F. Sonnen
Rachel A. Minerath, Rajesh K. Kasam, Casey O. Swoboda, Vikram Prasad, Kelly M. Grimes, N. Scott Blair, Hadi Khalil, Christina M Alfieri, Logan Eads, Anthony J. Saviola, Mohamad Azhar, Lianjie Miao, Mingfu Wu, Michelle Tallquist, Kirk C. Hansen, Matthew T Weirauch, Katherine E. Yutzey, Douglas P. Millay, Jeffery D. Molkentin
Márta Korbonits, Xian Wang, Sayka Barry, Chung Thong Lim, Oniz Suleyman, Stefano De Tito, Nazia Uddin, Maria Lillina Vignola, Charlotte Hall, Laura Perna, J. Paul Chapple, Gabor Czibik, Sian M Henson, Valle Morales, Katiuscia Bianchi, Viðar Örn Eðvarðsson, Kristján Ari Ragnarsson, Viktoría Eir Kristinsdóttir, Anne Debeer, Yoeri Sleyp, Rena Zinchenko, Glenn Anderson, Michael Duchen, Kritarth Singh, Chih Yao Chung, Yu Yuan, Sandip Patel, Artem O. Borovikov, Hans Tómas Björnsson, Hilde Van Esch, Sharon Tooze, Ezra Aksoy, Caroline Brennan, Oliver Haworth
Zukai Liu, Chengxiang Qiu, Connor A. Kubo, Stella Xu, Riza M. Daza, Eva Nichols, Wei Yang, Anh Vo, Mary B. O’Neill, Choli Lee, Jay Shendure, Nobuhiko Hamazaki
Kaleb Hill, Aaron H. Griffing, Michael A. Palmer, Bezia Lemma, Aria Lupo, Tony Gamble, Natalia A. Shylo, Andrej Košmrlj, Paul A. Trainor, Celeste M. Nelson
Olga D. Jarosińska, Amalia Riga, Hala Zahreddine Fahs, Joren M. Woeltjes, Ruben Schmidt, Fathima S. Refai, Suma Gopinadhan, Kristin C. Gunsalus, Mike Boxem
Sarah N. Steiner, Eric Horst, Mitre Athaiya, Craig N. Johnson, Joseph Y. Shen, Michelle L. Kerns, Geeta Mehta, Ramiro Iglesias-Bartolome, Pierre A. Coulombe
Akshai Janardhana Kurup, Aya Mikdache, Patricia Diabangouaya, Gwendoline Gros, Camila Garcia-Baudino, Cristian A. Undurraga, Andres F. Sarrazin, Pedro P. Hernandez
Rafael Casado-Navarro, Ana Bermejo-Santos, Rodrigo Torrillas-de la Cal, María Pilar Madrigal, Virgilia Olivé, Li Ying Chen-Chen, Sonia Amorós-Bru, Sandra Jurado, Esther Serrano-Saiz
Judhajeet Ray, Evelyn Jagoda, Maya U. Sheth, James Galante, Dulguun Amgalan, Andreas R. Gschwind, Chad J. Munger, Jacob Huang, Glen Munson, Madeleine Murphy, Eugenio Mattei, Timothy Barry, Vasundhara Singh, Aarthee Baskaran, Helen Kang, Eugene Katsevich, Lars M. Steinmetz, Jesse M. Engreitz
Ye Lynne Kim, Young-Woo Jo, Takwon Yoo, Kyusang Yoo, Ji-Hoon Kim, Myungsun Park, In-Wook Song, Hyun Kim, Yea-Eun Kim, Sang-Hyeon Hann, Jong-Eun Park, Daehyun Baek, Young-Yun Kong
Michael C. Mazzola, Ting Zhao, Anna Kiem, Trine A. Kristiansen, Karin Gustafsson, Lai Ping Wong, Emily Scott-Solomon, Marissa D. Fahlberg, Sarah Forward, Emane Rose Assita, Giulia Schiroli, Maris Handley, Youmna Kfoury, Tsuyoshi Fukushima, Samuel Keyes, Azeem Sharda, Jelena Milosevic, Hiroki Kato, Pavel Ivanov, David B. Sykes, Sheldon J. J. Kwok, Ruslan I Sadreyev, Vijay G. Sankaran, Ya-Chieh Hsu, David T. Scadden
Octavia Santis Larrain, Alice Alhaj Kadour, Sobhika Agarwala, Wantong Li, Bradley W. Blaser, Michael R. Lasarev, Roxana Alexandridis, Anthony Veltri, Khaliun Enkhbayar, Elliott J. Hagedorn, Owen J. Tamplin
Katharina Lemberg, Gijs A.C. Franken, Korbinian M. Riedhammer, Selina Hölzel, Kirollos Yousef, Kraisoon Lomjansook, Gina Kalkar, Caroline M. Kolvenbach, Daniel Marchuk, Elena Zion, Ken Saida, Florian Buerger, Friedhelm Hildebrandt
Declan J. Gainer, Kassandra M. Coyle, Matthew T. Rätsep, Douglas Quilty, Brian Tran, Sofia Skebo, M. Martin VandenBroek, Kimberly L. Laverty, Yupu Deng, Shawyon P. Shirazi, Hugh JM Brady, Jennifer M.S. Sucre, Eric Vivier, Niraj Shrestha, Hing C. Wong, Duncan J Stewart, Nicolle J. Dominik, Mark L. Ormiston
Wentao Han, Hassan Bjeije, Hamza Celik, Michael Rettig, Nancy Issa, Andrew L. Young, Yanan Li, Infencia Xavier Raj, Christine R. Zhang, Aishwarya Krishnan, Tyler M. Parsons, Samantha C. Burkart, Jason Arand, Wei Yang, Jeffrey A. Magee, Grant A. Challen
Michelle Lohbihler, Amos A. Lim, Stéphane Massé, Maggie Kwan, Omar Mourad, Olya Mastikhina, Brandon M. Murareanu, Malak Elbatarny, Renu Sarao, Beiping Qiang, Wahiba Dhahri, Matthew L. Chang, Alice L.Y. Xu, Amine Mazine, Shahryar Khattak, Sara S. Nunes, Kumaraswamy Nanthakumar, Michael A. Laflamme, Stephanie Protze
Cory P. Johnson, Hannah M. Somers, Sophie E. Craig, Heath Fuqua, Lynne Beverly-Staggs, Kailee E. Tanaka, Sydney M. Brown, Charles H. Toulmin, Matthew D. Cox, Joel H. Graber, Melissa S. Maginnis, Hermann Haller
Giorgio Anselmi, Vincent Frontera, Christina Rode, Andrew Jarratt, Naeema T. Mehmood, Matthew Nicholls, Stella Antoniou, Emanuele Azzoni, John Stamatoyannopoulos, Ditsa Levanon, Yoram Groner, Marella F.T.R. de Bruijn
İsmail Küçükaylak, Francisco Javier Martínez Morcillo, Kai Halwas, Nils Reiche, Manuel Metzger, Petra Comelli, Jürgen Brinckmann, Sabine Eming, Matthias Hammerschmidt
Meryam Beniazza, Masahito Yoshihara, Daniel F Kaemena, James Ashmore, Suling Zhao, Michael O’Dwyer, Emil Andersson, Victor Olariu, Shintaro Katayama, Abdenour Soufi, Kosuke Yusa, Keisuke Kaji
Yuancheng Ryan Lu, James C. Cameron, Yan Hu, Han Shen, Shintaro Shirahama, Alexander Tyshkovskiy, Zhaoyi Chen, Jiahe Ai, Daniel Y. Zhu, Margarete M. Karg, Lindsey A. Chew, George W. Bell, Siddhartha G. Jena, Yue He, Philip Seifert, Daisy Y. Shu, Mohamed A. EI-Brolosy, Qiannuo Lou, Bohan Zhang, Anna M. Puszynska, Xiaojie Qiu, Xiao Tian, Meredith Gregory-Ksander, Vadim N. Gladyshev, David A. Sinclair, Magali Saint-Geniez, Jason D. Buenrostro, Catherine Bowes Rickman, Bruce R. Ksander, Jonathan S. Weissman
Tom Levy, Chiara Anselmi, Katherine J. Ishizuka, Tal Gordon, Yotam Voskoboynik, Erin McGeever, Angela M. Detweiler, Liron Levin, Karla J. Palmeri, Daniel Dan Liu, Rahul Sinha, Benjamin F. Ohene-Gambill, Tal Raveh, Maurizio Morri, Virginia Vanni, Lucia Manni, Debashis Sahoo, Norma F. Neff, Benyamin Rosental, Irving L. Weissman, Ayelet Voskoboynik
Julia Schwarzpaul, Clara M. Droell, Afsheen Kumar, Harishny Sarma, Madeleine Gruenauer, Selen Z. Ucar, Julio C. Hechavarría, Andreas G. Chiocchetti, Denise Haslinger
Ioanna Peraki, Ioannis K. Deligiannis, Dimitris Botskaris, Marianna Stagaki, Haroula Kontaki, Elena Deligianni, Giannis Giannoulakis, Matthieu D. Lavigne, Celia P. Martinez-Jimenez, Iannis Talianidis
Eva-Sophie Wallner, Natalie Edelbacher, Liam Dolan
Supergene control of chiral development in mirror-image flowers Haoran Xue, Marco Saltini, Nicola Illing, Kelly Shepherd, Olivia Page-Macdonald, Oliver Marketos, Caroline Robertson, Anand Shankar, Sarah Süß, Christian Kappel, Saleh Alseekh, Eva E. Deinum, Robert A. Ingle, Michael Lenhard
Sjoerd Woudenberg, Andrew R.G. Plackett, Zhaodong Hao, Hidemasa Suzuki, Luis Alonso Baez, Cecilia Borassi, Thorsten Hamann, Minako Ueda, Jane A. Langdale, Joris Sprakel, Jasper van der Gucht, Dolf Weijers
Gabriele Panicucci, Vinay Shukla, Viktoriia Voloboeva, Leonardo Jo, Kees van Kollenburg, Sara Buti, Laura Dalle Carbonare, Francesco Licausi, Daan A. Weits
Bonnie K. Kircher, Antonia Weberling, Erin J. Vance, Natalia A. Shylo, Katherine Starr, Zoe B. Griffin, Hannah Wilson, Melainia McClain, Florian Hollfelder, Suzannah A. Williams, Thomas J. Sanger, Richard R. Behringer, Paul A. Trainor
Axel H Newton, Ella R Farley, Andrew T Major, Jennifer C Hutchison, Ben M Lawrence, Karen E Sears, Marilyn B Renfree, Aiden M C Couzens, Geoff Shaw, Sara Ord, Richard A Schneider, Andrew J Pask
Jasmine D. Alqassar, Teomie S. Rivera-Miranda, Joseph J. Hanly, Christopher R. Day, Silvia M. Planas Soto-Navarro, Paul B. Frandsen, Riccardo Papa, Arnaud Martin
Atsushi Saito, Stephanie Tankou, Kazuhiro Ishii, Makiko Sakao-Suzuki, Edwin C. Oh, Hannah Murdoch, Ho Namkung, Sunday Adelakun, Keiko Furukori, Masahiro Fujimuro, Paolo Salomoni, Gerd G. Maul, Gary S. Hayward, Qiyi Tang, Robert H. Yolken, Miles D. Houslay, Nicholas Katsanis, Isao Kosugi, Kun Yang, Atsushi Kamiya, Koko Ishizuka, Akira Sawa
Gonzalo Herranz, Diego Alonso-Larre, Tamara González, Laura Akintche, Alejandra Ramos-Manzano, Marta Iborra-Pernichi, María Velasco de la Esperanza, Covadonga Díaz-Díaz, Ian G Ganley, Patricia Boya, Sara Cogliati, Nuria Martínez-Martín, Fernando Martín-Belmonte
Daniel A. Reed, Anna E. Howell, Nadia Kuepper, Alice M. H. Tran, Astrid Magenau, Deborah S. Barkauskas, Max Nobis, Cecilia R. Chambers, Victoria Lee, Lily M. Channon, Jessie Zhu, Shona Ritchie, Janett Stoehr, Kaitlin Wylie, Julia Chen, Denise Attwater, Kate Harvey, Sunny Z. Wu, Kate Saw, Ruth J. Lyons, Anaiis Zaratzian, Michael Tayao, Andrew Da Silva, David Gallego-Ortega, Anthony J. Gill, Thomas R. Cox, Brooke A. Pereira, Kendelle J. Murphy, Jennifer P. Morton, Elgene Lim, Alexander Swarbrick, Sandra O’Toole, Michael S. Samuel, C. Elizabeth Caldon, Alexandra Zanin-Zhorov, Paul Timpson, David Herrmann
Rianne G. Bouma, Willem-Jan de Leeuw, Aru Z. Wang, Muddassir Malik, Joeke G.C. Stolwijk, Veronique A.L. Konijn, Anne Mensink, Natalie Proost, Maarten K. Nijen Twilhaar, Tibor van Welsem, Negisa Seyed Toutounchi, Alsya J. Affandi, Jip T. van Dinter, Fred van Leeuwen, Joke M.M. den Haan
Ronan Bouzignac, Amandine Palandri, Amal Zine El Aabidine, Thomas Mangeat, Tatiana Merle, Martine Cazales, Antonio Trullo, Christian Rouviere, Virginia Pimmett, Mounia Lagha, Magali Suzanne
Kiara C. Eldred, Matthew Wooten, Derek H. Janssens, Joshua Hahn, Shane J Neph, Sierra J. Edgerton, Gracious Wyatt-Draher, Stephanie M. Sherman, Jane E. Ranchalis, Andrew B. Stergachis, Thomas A. Reh, Steven Henikoff
Ryan G. Savill, Alba Villaronga-Luque, Marc Trani Bustos, Yonit Maroudas-Sacks, Julia Batki, Alexander Meissner, Allyson Q. Ryan, Carl D. Modes, Otger Campàs, Jesse V Veenvliet
Namwook Kim, Christine S. Kim, Junhee Park, Hyeon Jun Yoon, Ju Ang Kim, Yong Gun Kim, Joonhyung Kim, Injun Song, Donghyuck Ahn, Jihyeon Myeong, Byungmoo Oh, Jaeyoon You, Eunju Hong, Sukin Jeong, Kyungmoo Yea, Sung Won Kim, Ok Sarah Shin, Seung Joon Kim, Minho Lee, Myungin Baek, Youngtae Jeong
Jingyu Wang, Danail Stoychev, Mick Phillips, David Miguel Susano Pinto, Richard M. Parton, Nick Hall, Josh Titlow, Ana Rita Faria, Matthew Wincott, Dalia Gala, Andreas Gerondopoulos, Niloufer Irani, Ian Dobbie, Lothar Schermelleh, Martin Booth, Ilan Davis
Amine Mazine, Alexander A. Mikryukov, Ian Fernandes, Clifford Z. Liu, Soheil Jahangiri, Marcy Martin, Eric K. N. Gähwiler, Melanie Generali, Juliana Gomez, Neda Latifi, Yifei Miao, Yu Liu, Michael A Laflamme, Craig A Simmons, Simon P. Hoerstrup, Maximilian Y. Emmert, Bruce D. Gelb, Mingxia Gu, Gordon M. Keller
Lucien Hinderling, Hannah S. Heil, Alfredo Rates, Philipp Seidel, Manuel Gunkel, Benedict Diederich, Thomas Guilbert, Rémy Torro, Otmane Bouchareb, Claire Demeautis, Célia Martin, Scott Brooks, Evangelos Sisamakis, Erwan Grandgirard, Jerome Mutterer, Harrison Oatman, Jared Toettcher, Andrii Rogov, Nelda Antonovaite, Karl Johansson, Johannes K. Ahnlinde, Oscar André, Philip Nordenfelt, Pontus Nordenfelt, Claudia Pfander, Jürgen Reymann, Talley Lambert, Marco R. Cosenza, Jan O. Korbel, Rainer Pepperkok, Lukas C. Kapitein, Olivier Pertz, Nils Norlin, Aliaksandr Halavatyi, Rafael Camacho
Jack Nunn, Håkon da Silva Hyldmo, Lauren McKnight, Heather McCulloch, Jennifer Lavers, Julie Old, Laura Smith, Nicola Grobler, Cheryl Tan Kay Yin, Wing Yan Chan, Candice Raeburn, Nittya S. M. Simard, Adam Kingsley Smith, Sam Van Holsbeeck, Eleanor Drinkwater, Kit Prendergast, Emma Burrows, Christopher L. Lawson
 (3 votes)
(3 votes) (No Ratings Yet)
(No Ratings Yet)