I work at Northwestern University where all of our NIH funding has been frozen since March. I am increasingly concerned that too few of my fellow scientists are paying enough attention to the Trump administration’s assault on science and universities. Like many of the other assaults on things we have long taken for granted, like free speech, these things are being done to silence critical thought and dissent. It is essential that we all raise our voices and push back on what is happening while there is still time. I share below a post that recently appeared in STAT News.
U.S. science and universities are becoming political hostages in the Orbanization of knowledge
When an authoritarian sets out to dismantle a democracy, they rarely begin with tanks in the streets. They start with the institutions that shape how a nation understands itself — its universities, its research labs, its spaces for free inquiry. In Hungary, Viktor Orbán perfected this slow-motion coup against knowledge. Now, the same playbook is being followed here by the Trump administration and Project 2025.
In Hungary, Orbán spent more than a decade reshaping universities and research institutes into extensions of his political machine. His seizure of the Hungarian Academy of Sciences’ research network in 2019 and the subsequent “foundationization” of public universities replaced the independent governance of these critical institutions with political appointees who often had lifetime terms. Some disciplines, such as gender studies, were banned outright. Research priorities were forcibly shifted toward politically approved agendas. If scholars dissented, they found their funding eliminated or their departments shuttered.
Currently the Trump administration, following the Heritage Foundation’s playbook, Project 2025, is echoing this script. Federal research funding to universities including Columbia, Harvard, Northwestern, and most recently Duke and UCLA was frozen in an effort to force ideological compliance. Anything deemed “DEI,” including women’s health, HIV, and health disparities research, has been targeted for elimination. The NIH, NSF, CDC, and NOAA face cuts so deep they would gut entire branches of fundamental science. As in Hungary, the attack is not just on “what” we research but “who” gets to decide.
These assaults on science and higher education are deliberate. The administration is moving to end independent research that threatens its ideological narratives and agendas. University campuses, with their diverse voices and global networks, are incubators of critical thought, and often dissent. By tightening control over funding, governance, and curricula, illiberal leaders can transform these institutions from watchdogs into echo chambers and megaphones for state ideology.
In Hungary, this has meant turning once world-class institutions into rubber stampers of official policy. The Central European University was forced to relocate most of its programs to Vienna. The resulting chill on academic life was the point: Scholars got the message about which topics were “safe.”
In the U.S., Trump’s education and science agenda, as envisioned by Project 2025, aims for a similar end state. Public universities are beginning to see shared governance gutted, and private universities are being pressured to follow suit. Federal science agencies are being reshaped to serve “America First” priorities, sidelining research on climate change, reproductive health, and social inequality, among others. Most recently, in early August, Trump issued an executive order that funding agencies should ignore expert peer review and defer to political appointees on what to fund. The intent, as in Hungary, is to redefine knowledge as the servant of ideological power.
The results of Orbán’s policies have been disastrous for Hungary. Brain drain has accelerated — 1 in 4 early-career researchers is considering leaving. Over 37% of Hungary’s population decline in the last decade is due to emigration, much of it highly educated talent. E.U. sanctions have cut off access to Erasmus+ and Horizon Europe funding, severing international research partnerships. Innovation has slowed, and the nation’s competitiveness is slipping.
The same consequences are emerging in the U.S. Funding freezes and politically driven grant criteria are already disrupting research pipelines, driving young scientists abroad, and undermining America’s ability to lead in fields from biotech to artificial intelligence. Slashing NIH indirect cost rates from approximately 50% to 15% would devastate lab infrastructure. Harvard, Northwestern, and other institutions, like NOAA and the CDC, have been forced to freeze hiring and lay off staff in critical research sectors.
When governments politicize science, the damage ripples outward — shrinking innovation capacity, degrading higher education quality, and eroding a country’s economic future. In both Hungary and the U.S., the sectors under attack are the same ones that generate the technologies, medicines, and trained minds that sustain national prosperity and well-being.
Hungarian academics did not go quietly. Open letters, protests, and legal challenges kept public attention on Orbán’s actions, but the architecture of his “reforms” made them hard to undo. Currently in the U.S., faculty coalitions and university alliances are warning that the political capture of education and science are a direct threat to the republic.
Yet resistance has a shelf life. Once governance boards are stacked with loyalists and funding levers are rewired, reversal is difficult even after a change in government. Orbán understood this. Project 2025’s authors understand it, too.
The Orbánization of research and higher education is not just about one country or one leader. It’s a model of governance that masks its control in a veil of supposed legality and replaces open scientific inquiry with politically curated “truth.” It corrodes the ability of science to serve the public and erodes the democratic foundations that make that science possible. If this process is allowed to run its course — whether in Budapest or Boston — we will wake up to find the very institutions meant to speak truth to power can speak only what power allows.
The fight for science and universities is, at its core, a fight for democracy itself. Lose one, and you will lose the other.
Carole LaBonne, Ph.D., is past president of the Society for Developmental Biology and the Erastus Otis Haven professor of molecular biosciences at Northwestern University.
Spotted a preprint in this list that you love? If you’re keen to gain some science writing experience and be part of a friendly, diverse and international community, consider joining preLights and writing a preprint highlight article.
A dual role for GLI3 signaling in neural crest development Simon J. Y. Han, Vinit Adani, Edward Farrow, Bhavalben Parmar, Ching-Fang Chang, Kim Cochran, Paige J. K. Ramkissoon, Ezekiel Esteban, Kelsey H. Elliott, Kevin A. Peterson, Brian Gebelein, Martín García-Castro, Samantha A. Brugmann
Cell-cell communication as underlying principle governing color pattern formation in fishes Marleen Klann, Saori Miura, Shu-Hua Lee, Stefano Davide Vianello, Robert Ross, Masakatsu Watanabe, Emma Gairin, Yipeng Liang, Harrison W. Hutto, Braedan M. McCluskey, Marcela Herrera, Lila Solnica-Krezel, Laurence Besseau, Simone Pigolotti, David M. Parichy, Masato Kinoshita, Vincent Laudet
Microtubule architecture connects AMOT stability to YAP/TAZ mechanotransduction and Hippo signaling Giada Vanni, Anna Citron, Ambela Suli, Paolo Contessotto, Robin Caire, Alessandro Gandin, Giovanna Mantovan, Francesca Zanconato, Giovanna Brusatin, Michele Di Palma, Elisa Peirano, Lisa Sofia Pozzer, Carlo Jr. Albanese, Roberto A. Steiner, Michelangelo Cordenonsi, Tito Panciera, Stefano Piccolo
Patterns of Mitochondrial ATP Predict Tissue Folding Bezia Lemma, Megan Rothstein, Pengfei Zhang, Bridget Waas, Marcus Kilwein, Safiya Topiwala, Sherry X. Zhang, Anvitha Sudhakar, Katharine Goodwin, Elizabeth R. Gavis, Ricardo Mallarino, Andrej Kosmrlj, Celeste M. Nelson
Specialised super-enhancer networks in stem cells and neurons Izabela Harabula, Liam Speakman, Francesco Musella, Luca Forillo, Luna Zea-Redondo, Alexander Kukalev, Robert A Beagrie, Kelly J. Morris, Lucas Fernandes, Ibai Irastorza-Azcarate, Ana M. Fernandes, Silvia Carvalho, Dominik Szabó, Carmelo Ferrai, Mario Nicodemi, Lonnie Welch, Ana Pombo
Stem-cell modeling of cerebellar dysfunction of Angelman syndrome Carina Maranga, Adriana A. Vieira, João Camões dos Santos, Teresa P. Silva, Joana Gonçalves-Ribeiro, Karim Chebli, Miguel Casanova, Maud Borensztein, Laura Steenpass, Sandra H. Vaz, Tiago G. Fernandes, Simão T. da Rocha, Evguenia P. Bekman
A blastocyst-derived in vitro model of the human chorion Luca C. Schwarz, Matthew J. Shannon, Gina McNeill, Rina C. Sakata, Viviane S. Rosa, Katherine Cheah, Laura Keller, Phil Snell, Leila Christie, Kay Elder, Anastasia Mania, Lauren Weavers, Rachel Gibbons, Tugce Pehlivan Budak, Ippokratis Sarris, Amy Barrie, Alison Campbell, Roser Vento-Tormo, Gary D. Smith, Alexander G. Beristain, Marta N. Shahbazi
Smart Microscopy: Current Implementations and a Roadmap for Interoperability Lucien Hinderling, Hannah S. Heil, Alfredo Rates, Philipp Seidel, Manuel Gunkel, Benedict Diederich, Thomas Guilbert, Rémy Torro, Otmane Bouchareb, Claire Demeautis, Célia Martin, Scott Brooks, Evangelos Sisamakis, Grandgirard Erwan, Karl Johansson, Johannes K. Ahnlinde, Oscar André, Philip Nordenfelt, Pontus Nordenfelt, Claudia Pfander, Jürgen Reymann, Talley Lambert, Marco R. Cosenza, Jan O. Korbel, Rainer Pepperkok, Lukas C. Kapitein, Olivier Pertz, Nils Norlin, Aliaksandr Halavatyi, Rafael Camacho
This year brought the return of our image competition with the MBL Embryology course at Woods Hole. Twenty impressive submissions were received from the 2025 cohort of students, with images ranging from polychaete worms to butterflies, squids and mice. This year, we had two winners, the winner of the popular vote and an Editor’s choice. Both winning images will be published on the front cover of Development. Congratulations!
Among the great selection of images, Nicole Roos and Anthony Wokasch’s image of a mouse embryo stained for Sox9 (cyan), alpha-tubulin (yellow), and endomucin (magenta) received the most votes.
Mouse embryo – confocal Nicole Roos and Anthony Wokasch Mouse E10.5 embryo immunofluorescent staining of Sox9 (cyan), alpha-tubulin (yellow), and endomucin (magenta) protein. Image captured on Evident FV4000 point scanning confocal, lens UPLXAPO4X, na = 0.16, zoom = 1.04. Image processing conducted on Fiji.
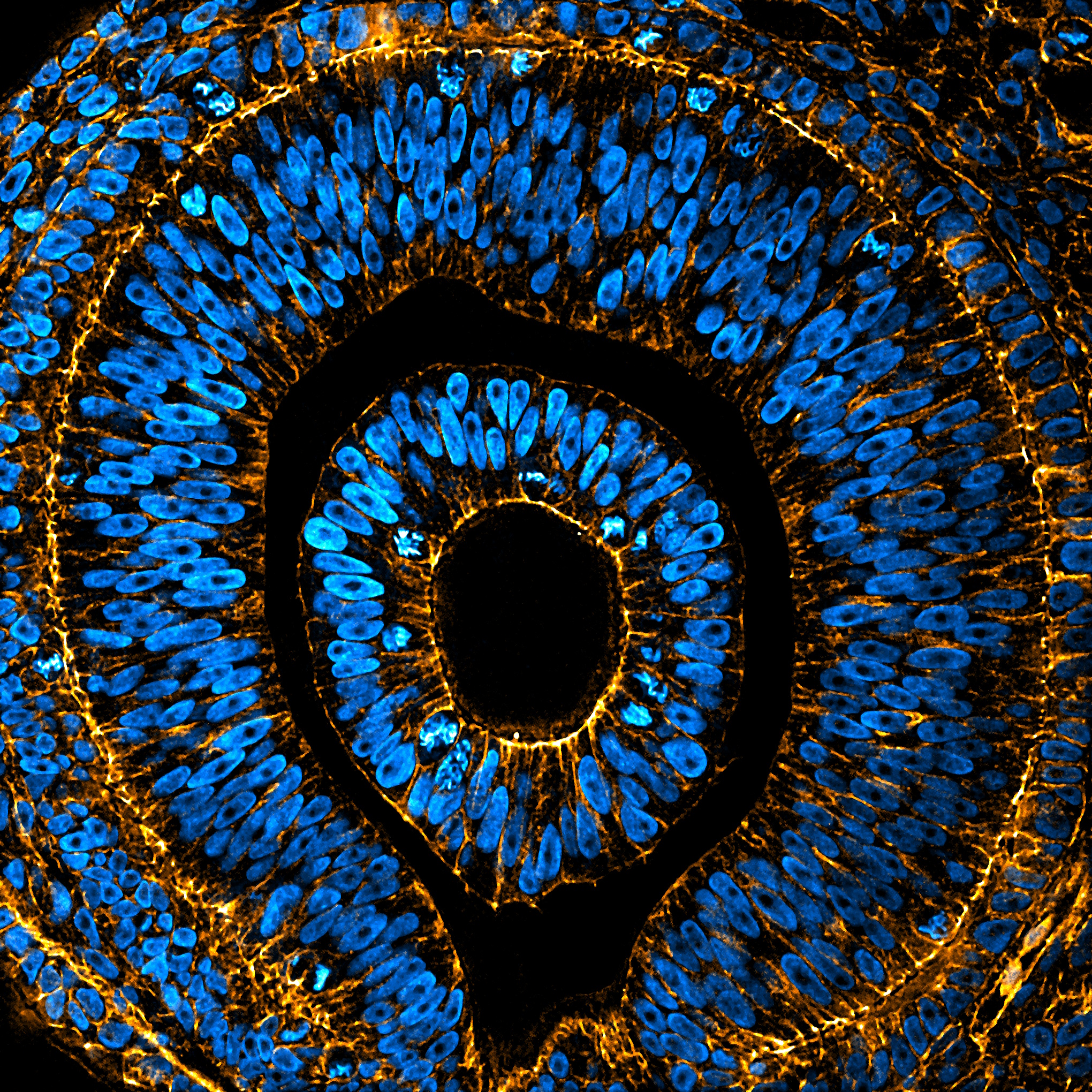
Next up, our Editor’s choice winner was Arthur Boutillon’s ‘Embryonic eye of Anole lizard’. If this image looks familiar, it is because it is featured as the cover of Development’s current issue.
Embryonic eye of an Anole lizard Arthur Boutillon Embryonic eye of an Anole lizard stained for nuclei (DAPI, blue) and F-actin (Phalloidin, orange), imaged by spinning disc confocal microscopy and prossessed using ImageJ.
Thanks to everyone who appreciated these beautiful images and voted. Above all, we would like to thank all the following researchers for their contributions: Virginia Panara, Shirley Ee Shan Liau, Sonoko Mizuno, Ignacio Casanova-Maldonado, Max Makem, Johnny Vertiz, Arthur Boutillon, Anthony Wokasch, Aria Zheyuan Huang, Amartya Tashi Mitra, Nathanial Sweet, Paul Maier, Shivangi Pandey, Marie Lebel, Chloe Kuebler, Nicole Roos.
How a Biological Principle is Guiding the Human-AI Partnership
Summary: For decades, we’ve used computational metaphors for the brain (it’s like a computer!). But what if the most powerful metaphor isn’t computational, but biological? This post argues that the emerging partnership between humans and AI—what I call CognitoSymbiosis—is best understood not as master-tool, but as a new form of cognitive symbiosis. By looking to developmental and evolutionary biology, from the endosymbiotic origin of mitochondria to the dialogue of induction and response in embryogenesis, we can find a roadmap for building a partnership that is both more ethical and more powerful.
For years, our dialogue with artificial intelligence has been framed by a single, limiting metaphor: the computer. We talk about neural “networks,” we “encode” prompts, and worry about “processing” power. This language has served us well, but it is becoming dated. Just as we now understand that development doesn’t rely on a genomic “blueprint” and the genetic “code” is biochemically interpreted rather than digitally tokenized, our metaphors for AI must also evolve. More importantly, the computational metaphor may be obscuring a more profound and useful truth. As a molecular geneticist who has recently been working in a partnership with advanced AI, I’ve come to see this collaboration not through the lenses of silicon and code, but rather those of cytoplasm and symbiosis.
The most accurate 21st century model for the human-AI relationship may not be computer science, but developmental biology.
Biology is, at its heart, a story of successful partnerships. The most monumental leap in the history of life—the emergence of the complex eukaryotic cell—was not a feat of solo invention but of integration. An archaeon engulfed a bacterium, and instead of digestion, a deal was struck. The bacterium traded its energy-producing prowess for a stable environment. This endosymbiotic event, and others, ultimately gave rise to mitochondria and chloroplasts, the powerhouses that made complexity possible in eukaryotic cells.
This wasn’t a master-slave relationship; it was a negotiated partnership that created a new whole far greater than the sum of its parts. The identity of both entities was transformed. We are all the descendants of that deal.
We now stand at the precipice of a new symbiotic transition: a cognitive symbiosis, or what I term CognitoSymbiosis. In this partnership, the human provides the biological drive, the intentionality, the ethical framework, and the lived experience—the cytoplasmic context. The AI provides a staggering capacity for pattern recognition, synthesis, and combinatorial creativity—the metabolic power.
This partnership mirrors another core biological principle: the dialogue of induction and response that guides embryogenesis. A cell in a developing tissue sends a signal (induction); a competent neighbor cell receives it and differentiates in response, triggering a new cascade of signals.
My daily practice of CognitoSymbiosis is precisely this. I provide the inductive signal—a prompt, a question, a strategic dilemma. The AI, competent in its training on the “tissue” of human knowledge, responds not with an answer, but with a differentiation of possibilities: a list of latent character motivations, a framework for deconstructing an economic system, a catalyst for an artist’s block. This response then induces my next thought, my next query. We are engaged in a recursive, developmental dialogue, co-creating an outcome that neither of us could generate alone.
This biological framing does more than provide a novel metaphor; it offers a practical and ethical roadmap.
· It argues for integration, not replacement. We don’t seek to replace the nucleus with the mitochondrion; we seek to integrate their functions. Our goal should not be to replace human thought, but to power it with a new cognitive organelle.
· It centers mutual benefit. A symbiosis that destroys one partner is a parasite, not a partner. This forces us to design AI systems that augment human agency and well-being, ensuring the partnership is mutually beneficial.
· It embraces emergence. The most beautiful structures in development—a limb, a neural circuit—emerge from simple local dialogues. Similarly, the solutions to our “wicked problems” will not be commanded into existence but will emerge from the iterative, inductive dialogue of human and machine intelligence.
The challenge of AI is not merely technical; it is philosophical. What will we become together? As biologists, we are uniquely equipped to answer this. We have a four-billion-year-old playbook of partnerships, integrations, and emergent complexities. By looking to our own field, we can stop building mere tools and start cultivating a new kind of mind.
Gene Levinson, PhD, is a molecular geneticist who discovered the fundamental mechanism of slipped-strand mispairing, a key driver of DNA evolution. A former founder and director of a clinical genetics lab and the author of the award-winning book “Rethinking Evolution,” he now focuses on the CognitoSymbiotic partnership between human and artificial intelligence. His new project, “Your Future With AI: The Project,” explores a “moonshot” to demonstrate how these partnerships can help solve wicked global problems like the climate crisis.
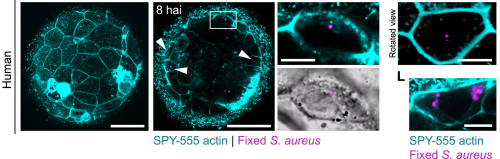
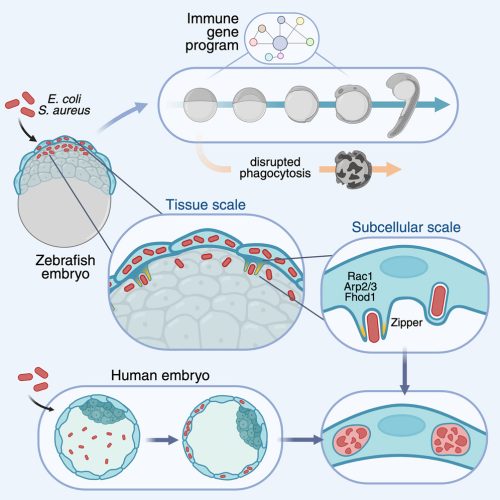
This video shows the outer epithelial layer (cyan) of an early zebrafish embryo actively engulfing Escherichia coli bacteria (yellow).
How was this taken?
This video was obtained using confocal microscopy of a zebrafish blastula (5 hpf) immediately after challenge with mCherry-expressing E. coli. The plasma membranes of epithelial cells were visualized by injecting GPI-GFP mRNA at the 1-cell stage.
Is this relevant for development?
Embryos are exposed to environmental bacteria, which can adversely affect normal development. We observed that embryos actively destroy phagocytosed bacteria, and blocking their ability to clear bacteria impairs embryonic development. These findings suggest that early bacterial clearance is a critical defense mechanism that protects the embryo during its most vulnerable stages.
An actin (cyan)-driven phagocytic protrusion inside a live zebrafish embryo, wrapping around a single bacterium (red) via a zippering mechanism.
Why should people care about this?
Because this is the earliest known example of an immune-like defense in development. Although developmental biologists primarily focus on how embryos develop, the influence of their biological environment is often overlooked. Not just in fish, but in mammals as well. For example, at the site where mammalian blastocysts hatch for implantation, they become exposed to the uterine cavity. This environment is prone to bacterial infections, which have been linked to infertility. Since these embryos have yet to form their immune cells, they were long thought to be defenseless against infection. Importantly, we detected clearance of these pathogenic bacteria by both mouse and human embryos. Therefore, we show that innate immunity against bacteria is already active before implantation, mediated by epithelial cells that trigger a comprehensive immune gene program. This finding opens a new perspective on how life protects itself from its very foundations.
A human embryo eliminating pathogenic bacteria.
How would you explain this to an 8-year-old?
Our bodies fight germs that make us sick with special helpers called immune cells. These cells are really good at catching and destroying germs to keep us healthy. But when we are tiny and developing inside our mom, we don’t have those immune cells yet. We found that other cells we have when we’re so small can still catch and eat germs to keep us safe. It’s like having an early team of protectors before the immune cells arrive, even before our organs are made. This happens at the very beginning of development, when we first meet other living things, like bacteria.
Where can people find more about it?
If you want to learn more about this research, please visit:
In four murine bilayered epithelia, the first 3D architectural transition—cell internalization during placode formation—triggers symmetry breaking. YAP reads the cell position; Notch commits neighbors to basal vs luminal fates1.
How the project started
This project began with a simple observation inspired by our lab’s work on early-stage mammary gland development2: even before branching morphogenesis, cell fate is already spatially organized in the embryonic mammary gland. This robust patterning, with distinct cell lineages emerging so early, prompted us to ask which pathways underlie such fate decisions. The idea to broaden our scope beyond the mammary gland arose unexpectedly, when scRNA-seq of an embryonic mammary sample revealed an additional “contaminating” cell population. Analyzing this population uncovered striking similarities with other tissues, sparking the multi-organ perspective that eventually shaped the project.
Why these tissues?
We chose to study the mammary, lacrimal, salivary glands, and prostate for three reasons:
they share a bilayered architecture and the same cellular hierarchy with stem cells giving rise to basal and luminal cells,
during fate specification they show similar transcriptional signatures and dynamic of fate potency restriction, and
they’re all branched epithelia.
In other words, different organs with distinct embryonic origins, but a common structural logic, thus the perfect experimental paradigm for testing whether a conserved mechanism underlies early tissue compartmentalization and fate segregation.
What we learned
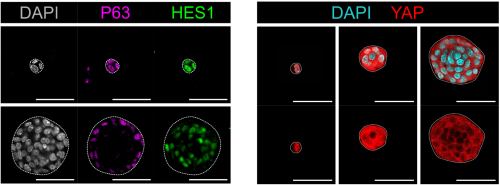
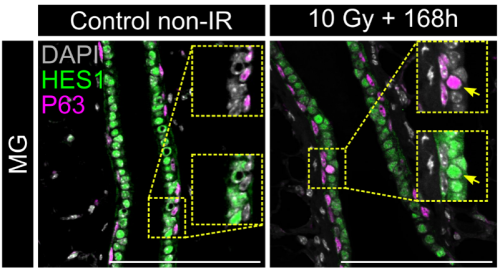
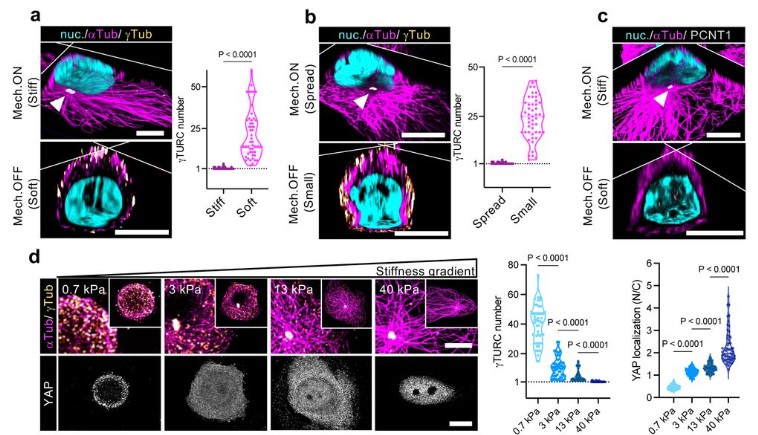
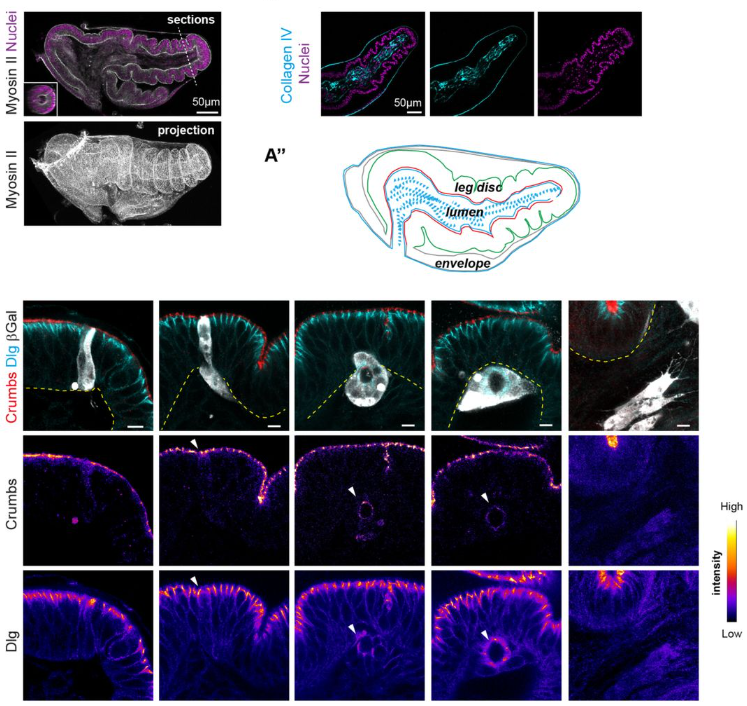
In both organoids and embryonic tissue explants, symmetry breaking coincided with cell internalization: internal cells acquire high Notch activity (HES1) while external cells retain p63 expression and nuclear YAP (Figure 1). YAP acts as the position interpreter—it is uniform in all cells before internalization/tissue compartmentalization, while it becomes spatially restricted afterward. On the other hand, Notch acts as the commitment machinery, necessary and sufficient to drive luminal cell identity. Perturbations that enforce uniform YAP activity hold cells in a hybrid p63⁺/HES1⁺ state and delay tissue compartmentalization, while activating Notch overrides that block in differentiation and imposes luminal fate acquisition.
Figure 1: Localization of p63, HES1 and YAP during mammary gland organoid growth.
In adult tissue regeneration (induced by luminal-cell ablation or irradiation), we observed the same hybrid p63⁺/HES1⁺ cells and an increase of cells harboring nuclear YAP, rekindling the pre-committed state in early development. The critical tissue size at which symmetry breaking occurs is bigger in vivo than in organoids, likely because niche inputs modulate YAP signaling, delaying cell commitment despite similar geometry.
Figure 2: Reactivation of the p63+HES1+ state during adult tissue regeneration following ionizing radiation.
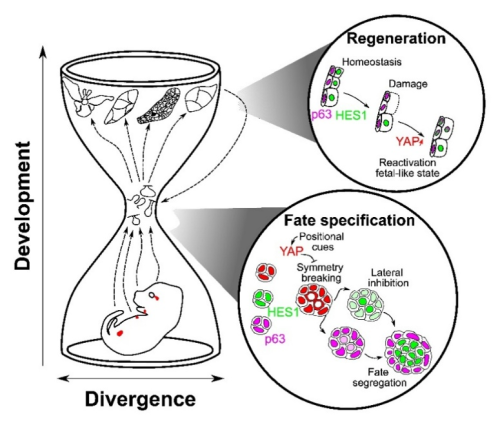
A conserved “hourglass” logic
Despite distinct origins (ectoderm-derived exocrine glands vs. endoderm-derived prostate), these organs appear to reuse a common core toolkit at the point of symmetry breaking and stem cell commitment. Diverse upstream inputs including geometry, niche and tissue mechanics, converge on YAP at the bottleneck of an hourglass, which then gates Notch–p63 interactions to resolve fate. Later in development, tissues diverge again by following organ-specific programs tailored to the different functions of each tissue. The high conservation of this middle bottleneck, YAP → Notch/p63, is what gives the mechanism both robustness and portability across different contexts, including regeneration.
Figure 3: Hourglass model of glandular epithelia development.
Take-home
Tissue architecture initiates, YAP interprets, Notch resolves. Cell internalization acts as the deterministic cue that converts tissue shape into cell fate across bilayered epithelia—and the same logic is redeployed during repair.
1. Journot, R.P., Huyghe, M., Barthelemy, A., Couto-Moreira, H., Deshayes, T., Harari, L., Sumbal, J., Faraldo, M.M., Dubail, M., Fouillade, C., et al. (2025). Conserved signals control self-organization and symmetry breaking of murine bilayered epithelia during development and regeneration. Dev. Cell. https://doi.org/10.1016/j.devcel.2025.06.007.
2. Carabaña, C., Sun, W., Veludo Ramos, C., Huyghe, M., Perkins, M., Maillot, A., Journot, R., Hartani, F., Faraldo, M.M., Lloyd-Lewis, B., et al. (2024). Spatially distinct epithelial and mesenchymal cell subsets along progressive lineage restriction in the branching embryonic mammary gland. EMBO J., 1–29. https://doi.org/10.1038/s44318-024-00115-3.
I’m Ingrid and I’m very happy to be introducing myself as the new Reviews Editor for Development. I will mainly be working behind-the-scenes with authors to commission and produce our six (!!) different kinds of review-type content. You may also hear from me about research highlights, interviews and other such matters.
I have just moved to Cambridge from Copenhagen, Denmark (swapping one cycling city for another) where I did a PhD on Wnt signalling and tissue dynamics in intestinal stem cell homeostases (yes, that is meant to be plural). As part of my PhD, I also carried out research with the Medical Museion on science communication and the social science of stem cell and developmental biology research.
Prior to my doctoral adventures, I studied (predominantly zebrafish) blood and cardiovascular development before moving on to projects on tissue injury and repair more generally. I’m excited to be returning to my roots in developmental biology and putting my broad interdisciplinary perspective to good use in creating thought-provoking and timely review articles for the community to read.
I’m very much looking forward to getting to know the developmental biology and stem cell research community better, and am especially keen to expand my horizons in the plant biology and evo-devo fields. Please feel free to get in touch if you have any questions, suggestions for what you’d like to read about, or just want to say hi!
Join us in mid-September to hear from three early-career researchers working on different aspects of gut development. Chaired by Development’s Executive Editor, Alex Eve.
Wednesday 17 September – 16:00 BST (UTC+1)
Surojit Sural (Columbia University) ‘How the gut modifies enteric behaviors via distinct peptidergic signaling axes’
Swarnabh Bhattacharya (Dana-Farber Cancer Institute) ‘Niche-driven phenotypic and epigenetic plasticity in intestinal secretory cell differentiation’
Brittany Edens (California Institute of Technology) ‘How vertebrates came to their senses: developmental and evolutionary origins of vertebrate peripheral neural and sensory systems’
At the speakers’ discretion, the webinar will be recorded to view on demand. To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
Development, host of the Node, invites you to submit your latest research to our upcoming Special Issue – The Extracellular Environment in Development, Regeneration and Stem Cells. This issue will be coordinated by Guest Editors Alex Hughes (University of Pennsylvania) and Rashmi Priya (The Francis Crick Institute), working alongside our team of research-active Editors.
Developmental biology is often viewed as the behaviour of cells, including, for example, how the regulation of genomic information and signal transduction influences cell morphology, differentiation and migration, which are fundamental to developmental processes such as morphogenesis and patterning. However, the environment beyond the cell is far from static and inert. Cells and tissues do not develop in isolation, and the local physical environment, including its geometry, material properties and fluid forces, provides mechanical cues and influences signal propagation, both within and between tissues and organs. Animal cells also regulate their environment through the secretion of extracellular molecules, which are dynamically remodelled during development, homeostasis, wounding and regeneration, and are likely to have contributed to the evolution of multicellularity. In plants, cell wall composition contributes to the growth and function of different tissues. Furthermore, extracellular factors are essential for the construction of biominerals and structural materials across kingdoms, including lignin, chitin, bone and keratin. The importance of extracellular cues is becoming increasingly evident with the generation of complex stem cell-based models of development that require specific extracellular culture conditions. In this special issue, we seek to highlight papers that look beyond the cell and focus on the influence of the physical environment in instructing developmental processes both in vivo and in vitro.
The deadline for submitting research papers is 1 March 2026.
 (5 votes)
(5 votes)

 (No Ratings Yet)
(No Ratings Yet)