Supervisors: Jill Harrison, Keith Edwards and Chris Burt (RAGT Seeds)
Project Description
Summary: Ensuring continuous global food security will be a major challenge of the 21st century, and wheat contributes approximately 20% of the total calories consumed by humans (FAO, 2017). In cereals like wheat, inflorescence (ear) size determines the number of flowers (florets) and grains produced, and this aspect of plant architecture is regulated by the activity of stem cells in the growing shoot tips. The CLAVATA peptide/ receptor-like kinase signalling pathway maintains the size of the stem cell pool during plant development, and mutants in maize and tomato have increased yields, arising due to an increase in size of the stem cell pool. This project aims to intercept wheat CLAVATA signalling to engineer ears with more fertile grain sites and increase yield.
The project will involve:
(1) Identification of wheat CLAVATA pathway components
(2) Expression analyses of wheat CLAVATA pathway components
(3) Generation phenotypic analysis of wheat CLAVATA pathway mutants.
Dr Harrison’s group has recently published gene trees for CLAVATA pathway components from a range of land plants (Whitewoods et al. (2018)), and she has experience of analysing gene expression patterns and function in a wide range of plant species. Professor Edwards and colleagues from the Bristol Centre for Agricultural Innovation have extensive experience with wheat having sequenced the genome (Brenchley et al. (2012)), identified many mutants from the exome sequenced Cadenza TILLING mutant population (Krasileva et al. (2017)) and established engineering procedures using CRISPR/Cas9. The CASE partnership with RAGT seeds will bring an opportunity for the student to directly experience wheat breeding and exchange knowledge and findings with wheat growers.
Training: By combining computational and wet lab approaches, your project work will provide training at the cutting edge of the plant development field. You will benefit from further formal teaching and internships included in the SWBioDTP programme. The skills and techniques you learn will be broadly applicable in the academic biology and biotech sectors and widely transferable amongst areas such as science policy, publishing and computing.
Funding Notes
A fully-funded four year SWBio DTP studentship will cover:
– a stipend* (at the standard UKRI rate; £15,009 per annum for 2019-2020)
– research and training costs
– tuition fees (at the standard UKRI rate)
– additional funds to support fieldwork, conferences and a 3-month internship
This is a CASE DTP studentship. As part of the programme, you will be required to undertake a placement with the CASE partner for a minimum of 3 months.
References
Brenchley et al. (2012). Analysis of the bread wheat genome using whole-genome shotgun sequencing. Nature 491: 705-710. Food and Agriculture Organization of the United Nations, FAOSTAT statistics database, Food balance sheets (2017); www.fao.org/faostat/en/#data/FBS. Krasileva et al. (2017). Uncovering hidden variation in polyploid wheat. PNAS 114: E913-E921. Whitewoods et al. 2018. CLAVATA was a genetic novelty for the morphological innovation of 3D growth in land plants. Current Biology 28: 2365-2376.
PhD position available at Liverpool John Moores University, starting Feb 2020. Extraction of exosomes from maternal blood for analysis of miRNA cargo and glycoprotein epitopes.
The Poulain lab (www.poulainlab.org) at the University of South Carolina in Columbia, SC seeks talented and motivated postdocs! Weuse zebrafish as a vertebrate model system and a unique combination of genetic, embryological and live imaging approaches to study the cellular and molecular mechanisms of axon guidance and degeneration during the formation of neural circuits in vivo. Specific projects include single-cell topographic transcriptomics and the formation of topographic maps, role of neural activity in topographic map plasticity, role of cell adhesion in axon developmental degeneration, and spatio-temporal control of trans-axonal degeneration signaling.
Candidates should hold a PhD in neurobiology or neuroscience and have a strong interest in neural development. Significant experience in molecular biology, genetics and fluorescence imaging approaches is required. Experience with zebrafish is desired but not mandatory. Interested candidates should email their resume/CV, the names and contact information of 3 references, and a cover letter explaining their interest to fpoulain@mailbox.sc.edu
NEUCrest Ph.D Studentships in Developmental Biology, Stem Cell Biology and Cancer Biology
Applications are invited from suitably qualified candidates for full-time fixed term positions as Early Stage Researchers in the lab of Prof Karen Liu, King’s College London (karen.liu@kcl.ac.uk), Centre for Craniofacial and Regenerative Biology, King’s College London.
These positions are funded by the Horizon 2020 programme of the European Union and will be available from 1st January 2020. Appointments will be on a full-time basis for a period of 3 years. Remuneration will be in line with the European Commission rules for Marie Skłodowska-Curie grant holders (Early-Stage Researchers, Initial Training Network).
NEUcrest is a four-year EU Horizon 2020 project funded by Agreement 860635. The neural crest is an essential stem cell population of vertebrate embryos. The project focuses on integrating academic, clinical and industrial research for a better understanding of neural crest development and neural crest related diseases. These pathologies are a major group of congenital diseases in human, and a heavy societal concern. The NEUcrest network comprises 20 partners in academia, industry and hospitals from seven European countries, gathered in a synergistic effort to advance knowledge and outreach of these diseases.
The Ph.D projects are highly multidisciplinary and will develop scientific strategies from experimental embryology, genome editing, imaging and generation of genomic datasets. A full list of projects and partners:
PhD Projects available through the lab of Prof Karen Liu, King’s College London:
PROJECT 1: The Role of ALK and GSK3 in normal and pathogenic neural crest migration. This project led by Prof Karen Liu (King’s College London) and Prof Angela Nieto (UMH, Spain). Applications see: https://tinyurl.com/yycpnlxj
Person Specification: First degree or Masters in Biological Sciences, Cell Biology, Genetics and Molecular Biology. Mobility requirement: EU applicants are eligible to apply. Applicants must not have been based in the country of registration for more than 12 months in the last 3 years.
Start date: Positions are available from 1st January 2020. Informal enquiries to Prof Karen Liu (karen.liu@kcl.ac.uk)
To Apply: Applications, in English, should include a detailed CV, certificates of examination grades (bachelor/master), a letter describing your career goals, skills and experience, as well as two letters of recommendation. Applications: https://tinyurl.com/yycpnlxj. Informal inquiries may be made to karen.liu@kcl.ac.uk. Deadline for applications is 15 November 2019, or until posts are filled.
PROJECT 2: Generation of neural crest cells from patient-derived hiPSCs for disease modelling and therapeutic applications. This industrial project will be led Dr Erin Knock and Dr Wing Chang with student based at STEMCell Technologies, Cambridge UK. Academic registration and supervision by the lab of Prof Karen Liu (King’s College London).
PROJECT 3: Establishment of human iPSCs from syndromic neurocristopathy patients with skeletal dysplasia This project is suitable for a clinical or suitably qualified non-clinical PhD student and is led by Prof Irene Mathijssen (email to i.mathijssen@erasmusmc.nl Erasmus Medical Centre, Netherlands) with secondment to Prof Karen Liu (King’s College London) and STEMCell Tech, Cambridge UK and Phenocell, France.
Cell adhesion molecules (CAMs) of the plasma membrane physically tie cells together into tissues, both via interaction of cells with other cells or with ECM (extracellular matrix); they also constitute hubs of information exchange with neighbouring cells and the environment. CAMs mediate a broad range of biological functions and their aberration contributes to a range of diseases, such as cancer progression or aberrant growth/regeneration of nerves.
On this project, you will study integrins, which are the major adhesion receptor class for extracellular matrices.Integrin signalling is performed by its adhesome, the dynamic population of proteins that link integrins both to the cytoskeleton and to signalling pathways via the cytoplasmic tails of the receptors. The composition and functional state of integrin-associated complexes essentially determines the adhesion state and signalling output.
To understand this, Martin Humphries has long-standing expertise in using proteomics based on mass spectrometry as a means to identify components of the integrin adhesome and study their function in mammalian cell culture [1]. On this project, you will extend such approaches to brain tissues in the living organism and study the function of identified proteins in the context of nerve growth.
For this, you will capitalise on the expertise of Andreas Prokop, who uses neurons of the fruit fly Drosophila to study mechanisms of nerve growth [2]. In this functional context, his team discovered and analysed important roles of integrins and some of their conventional adhesome components (new unpublished results). The small size and genetic tractability of Drosophila makes it feasible to perform proteomics on integrin adhesion complexes extracted from nerve cells of living organisms.
Once candidate components have been identified, you will then be able to capitalise on highly efficient genetic means provided by Drosophila to study their functional contributions during nerve growth [2] – potentially identifying promising drug targets for nerve regeneration therapies. A likely option will be the extension of such functional studies into mammalian models.
Your experimental skill training opportunities will include molecular biology, protein biochemistry, mass spectrometry, genetics, cell biology, a range of imaging techniques, bioinformatics, as well as insights into important concepts of the cell and neurobiology fields. In addition, AP is an expert in science communication providing further training opportunities important for your career development.
Epithelial tubes often have a functional polarity written along their proximo-distal (P-D) axis, with different segments of specialised cell-types carrying out distinct physiological activities. With a handful of notable exceptions, we know very little about how P-D axes and segment-specific differentiation are regulated during organogenesis.
The major objective of this project is to explore the molecular and cellular mechanisms that pattern and maintain functional polarity along the P-D axis in a structurally simple, but functionally sophisticated epithelial tube: the insect renal tubule. Approaches will include: state-of-the-art imaging, single cell RNAseq, genetics, physiological assays and in silico modelling.
The project will be carried out in the laboratory of Barry Denholm (Biomedical Sciences, University of Edinburgh)
A recent example of our work from a related project: Beaven, R. and Denholm, B (2018) Release and spread of Wingless is required to pattern the proximo-distal axis of Drosophila renal tubules eLife 2018;7:e35373 https://elifesciences.org/articles/35373
Miki Ebisuya’s research group at EMBL Barcelona is looking for a postdoctoral fellow who tackles a mystery in developmental biology: why larger animal species tend to show slower time. Why is the gestation period of humans ~9 months while that of mice is only 20 days? What are the molecular mechanisms of such inter-species timing differences?
Thanks to in vitro differentiation of iPS/ES cells, we can now recapitulate several developmental processes of diverse species, including humans. Thus, our plan is to systematically measure biochemical parameters, such as degradation rates, by using proteomics and RNAseq, and to compare them among species.
We are looking for a person who will carry out both cell biology experiments and high throughput data analyses (both can be taught by current lab members and collaborators). For more information, see the job description/application page.
* This is a position supported by EMBL internal grant as well as PGC2018-097872-A-I00 (MCIU/AEI/FEDER, UE) funded by the Spanish Ministry of Science, Innovation and Universities (MCIU) and co-funded by the European Regional Development Fund (ERDF, EU).
A Wellcome Trust/Royal Society funded Research Associate position is available in Dr. Kyra Campbell’s research group. This is a fantastic opportunity to join the Campbell group, who are focused on identifying the molecular mechanisms underlying epithelial cell plasticity during development and disease. We study this during morphogenesis of the Drosophila midgut (Campbell et al, Dev Cell 2011; Campbell and Casanova, Nat Comms 2015), and also in exciting Drosophila cancer models that we have recently generated (Campbell and Casanova, Plos Genetics 2018; Campbell et al, Nat Comms 2019).
We are combining single-cell OMICs approaches and deep-tissue imaging on our own labs dedicated multiphoton confocal microscope, with genetic approaches and CRISPR/Cas9 technologies. We are looking for a motivated and enthusiastic candidate who will play a central role in the lab. You must have a good honours degree and a PhD (or be close to completion) in areas relevant to cell/developmental biology (or have equivalent experience), along with experience in in vivo imaging and image analysis. Applicants are expected to have excellent interpersonal and communication skills, be highly independent and committed to research in a fast-moving and competitive field.
All cells in the body contain the same genetic material. The difference between cells therefore depends solely on which genes are expressed or ‘turned on’. Now, researchers from the University of Copenhagen have gained new insights into how genes are turned on and off and how the cells “forget their past” while developing into a specific cell in the body. This new knowledge is published in Nature and will be crucial for stem cell therapy and potentially treating people with cancer.
Stem cells all share the potential of developing into any specific cell in the body. Many researchers are therefore trying to answer the fundamental questions of what determines the cells’ developmental fate as well as when and why the cells lose the potential of developing into any cell.
Now, researchers from the Novo Nordisk Foundation Center for Stem Cell Biology (DanStem) at University of Copenhagen have discovered how stem cells can lose this potential and thus can be said to “forget their past”. It turns out that the proteins called transcription factors play another role than the scientists thought. For 30 years, the dogma has been that transcription factors are the engines of gene expression, triggering these changes by switching the genes on and off. However, new research results published in Nature reveal something quite different.
“We previously thought that transcription factors drive the process that determines whether a gene is expressed and subsequently translated into the corresponding protein. Our new results show that transcription factors may be more analogous to being the memory of the cell. As long as the transcription factors are connected to a gene, the gene can be read (turned on), but the external signals received by the cells seem to determine whether the gene is turned on or off. As soon as the transcription factors are gone, the cells can no longer return to their point of origin,” explains Josh Brickman, Professor and Group Leader, DanStem, University of Copenhagen.
The question of how a cell slowly develops from one state to another is key to understanding cell behavior in multicellular organisms. Stem cell researchers consider this vital, which is why they are constantly trying to refine techniques to develop the human body’s most basic cells into various specific types of cells that can be used, for example, to regenerate damaged tissue. So far, however, investigating the signals required to make cells switch identity has been extremely difficult, since making all the cells in a dish do the same thing at the same time is very difficult.
A protein centered viewpoint
The researchers developed a stem cell model to mimic a cell’s response to signaling and used it to, for first time, precisely determine the sequence of the events involved in a gene being turned on and off in response to a signal in stem cells. The researchers were able to describe how genes are turned on and off and under what circumstances a cell can develop in a certain direction but then elect to return to the starting-point. Part of this work involved measuring how proteins in a cell are modified by phosphorylation using advanced mass spectrometry available through an important collaboration with Jesper Olsen’s Group at the Novo Nordisk Foundation Center for Protein Research. “Combining forces with the Olsen group in the CPR enabled us to provide a unique deep description of how individual proteins in a cell react to signals from the outside,” continues Josh Brickman.
New answers to old scientific questions
These results are surprising. Although the sequence of cell transcription processes could not previously be measured as accurately as in this study, the dogma was that transcription factors comprise the on-off switch that is essential to initiate transcription of the individual gene. This is not so for embryonic stem cells and potentially for other cell types.
“Transcription factors are still a key signal, but they do not drive the process, as previously thought. Once they are there, the gene can be read, and they remain in place for a while after the gene is read. And when they are gone, the window in which the gene can be read can be closed again. You can compare it with the vapour trails you see in the sky when an airplane has passed. They linger for a while but slowly dissipate again,” explains first author, William Hamilton.
This discovery is first and foremost basic knowledge, which changes fundamental assumptions in molecular biology. The new results are especially important for researchers working on stem cells and cancer biology. They provide new insight into how cells develop, how pathways involved in development determine when cells change, and when the point of no return is reached. These pathways are also found frequently mutated in cancer and the findings in this study will be valuable to the study of malignant development.
“In the project, we focused on the fibroblast growth factor (FGF)–extracellular signal–regulated kinase (ERK) signalling pathway, which is a signalling pathway from a receptor on the surface of a cell to DNA inside the cell nucleus. This pathway is dysregulated in many types of cancer, and we therefore hope that many of the data in this study will help to inform aspects of cancer biology by indicating new ways to specifically target this signalling pathway in cancer cells,” concludes Josh Brickman.
They study was funded by the Novo Nordisk Foundation, the Independent Research Fund Denmark, the Danish National Research Foundation, the Human Frontier Science Program and the Lundbeck Foundation. It also involved an important collaboration with the group of Naama Barkai, at the Weizmann Institute for Science, Rehovot, Israel.
Novo Nordisk Foundation Center for Stem Cell Biology, DanStem, University of Copenhagen | joshua.brickman@sund.ku.dk
Josh Brickman has a background in molecular biology and gene regulation. From a PhD focused on transcriptional regulation he trained in developmental biology as a post-doctoral fellow, working in early mouse, and Xenopus, as well as cultivating embryonic stem cells as a model for developmental biology. He began his own lab with research projects bridging early development in multiple models systems with ES cells in a hybrid approach aimed at understanding conserved mechanisms of lineage specification, pluripotency and self-renewal. He currently seeks to understand how transcription factors regulate cell fate choice in ES cells and early embryos. More specifically, Professor Brickman’s and his group investigate the basis for transcriptional priming and commitment in ES cells and early in the specification of the endoderm lineage. They hope to understand the relevance of these molecular events to cellular decision making, pattern formation, in addition to stem and progenitor cell potency.
Novo Nordisk Foundation Center for Stem Cell Biology, DanStem, University of Copenhagen | william.hamilton@sund.ku.dk
William obtained his PhD at the Edinburgh University in the labs of Tilo Kunath and Mike Tyers, where he worked on defining factors that regulate MAPK signalling in mouse embryonic stem cells. He then joined the Brickman lab in Copenhagen where he expanded upon this to uncover how MAPK signalling regulates transcription and plasticity during early stem cell differentiation.
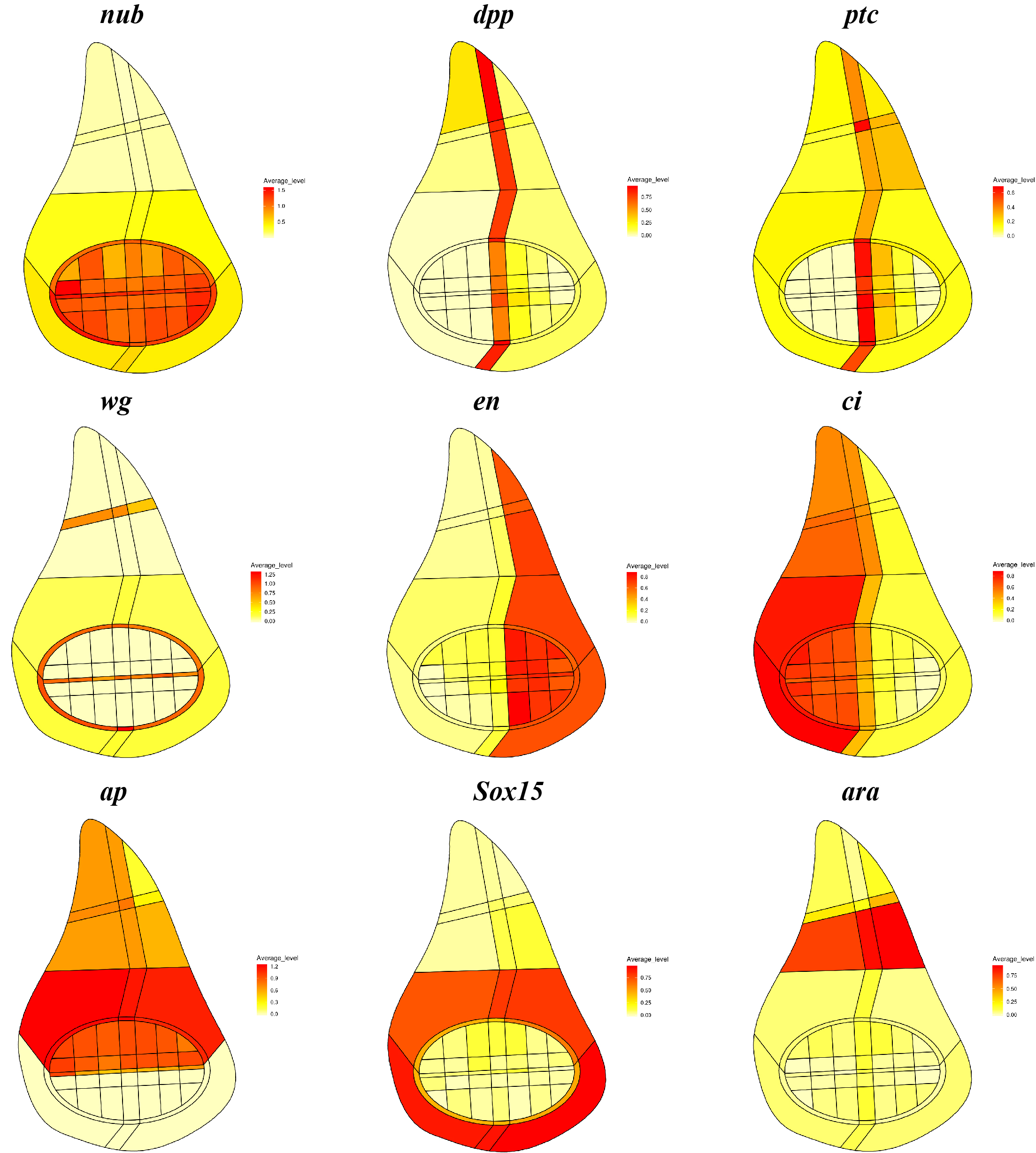
Drosophila wing discs are epithelial sac-like organs and a powerful model for investigating the link between proliferation and patterning. Of particular interest is the question of how single cells in the disc integrate information regarding position and growth control, as morphogens that pattern an axis can also regulate cell division. A new Techniques and Resources article in Development reports the application of single cell sequencing technologies to dissociated discs in an effort to understand these problems. We caught up with first author Mingxi Deng and his supervisor Yan Yan, Assistant Professor at The Hong Kong University of Science and Technology (HKUST), to hear more about the story.
Mingxi and Yan (L-R)
Yan, can you give us your scientific biography and the questions your lab is trying to answer?
YY My lab is primarily interested in organ size control, in particular, the roles of cell structural components such as apicobasal polarity proteins and cytoskeletal proteins in this process. When I was a graduate student with Prof. Trudi Schupbach at Princeton University, New Jersey, USA, I performed a genetic screen for mutants affecting Drosophila follicle cell epithelial morphogenesis and proliferation. I then got my postdoc training with Prof. Chris Doe at the University of Oregon, USA, where I learned how Drosophila embryonic neuroblasts lose their apical domains and emerge from neuroepithelia. These experiences were important for me to learn that cell polarity and, more broadly, cell structural proteins, are important for organ size and shape. Another important thing I learned from graduate school is the power of quantification, which influences how we approach questions now in the lab.
Mingxi, how did you come to work in the Yan lab, and what drives your research today?
MD When I was looking for a postgraduate student position at HKUST I found Prof. Yan’s research interesting in combining the power of Drosophila genetics with quantitative biology methods. Her lab also has a good reputation for being supportive to students, so I decided to join. I have always wanted to become a scientist and I get excited when I encounter new problems and need to find a way to solve them.
What was the drive behind doing a single cell analysis of the disc, and how easy was it to set the system up?
MD & YY It started with our study of how scribble (scrib) mutant tumours – which show disrupted tissue architecture – change over time. We found that they showed a high degree of plasticity, and suspected that they might be more heterogeneous than previously assumed. For this we needed to understand how much of the cell heterogeneity in the scrib tumours comes from heterogeneity already existing in wild-type wing discs. That was the starting point of this analysis.
We were lucky to have the help we needed for this study. Prof. Ting Xie from the Stowers Institute for Medical Research, Missouri, USA, happened to visit my university at the time. I went to talk with him and he kindly shared his unpublished fly single cell dissociation protocol. It is also very helpful to have colleagues Jiguang Wang and Hao Ge with whom to discuss methods: their expertise in bioinformatics and mathematics ensures that we are analysing data correctly and robustly. In addition, the single cell community has been very good in providing open-access analytical tools with user-friendly tutorials.
Can you give us the most surprising finding from your paper?
MD & YY The most surprising finding is that pattern formation partially persisted in the scrib mutant tumours. This is surprising because the morphogens important for pattern formation need to properly spread in space, and it suggests that further studies are needed to understand why particular pattern formation processes are robust against loss of tissue architecture.
A combination of different gene expression patterns from the ‘Virtual wing disc in situ’ section of the online database.
What does your scrib tumour model analysis tell you about the link between patterning and growth control in the disc?
MD & YY This is a very good question, but we still do not understand the link between patterning and growth control, although we are able to make more quantitative observations from these data. It was previously shown, with very clear genetic evidence, that patterning factors such as dpp are needed to ensure proper proliferation and growth in the disc. Previous experiments have also shown that cell proliferation and growth are more or less uniform throughout the whole disc. Now, we provide another line of evidence that proliferation and growth states do not appear to be particularly biased in sub-regions marked by any single patterning gene, in both wild-type and scrib mutant discs. Our data also suggest that a well-defined distribution of proliferation and growth states exists in discs and this distribution is severely disrupted in the scrib mutants. Interestingly, the temporal scrib mutant data suggested a positive correlation between formation of correct patterns and a distribution of proliferation and growth states closer to wild type.
Your single cell datasets are available to explore on a database: what questions do you think this database will be particularly useful for addressing?
MD & YY Wing discs have been a very good system to study pattern formation, organ size control and regeneration. I hope that our database can provide a good reference point for the community interested in these questions. For example, for researchers interested in how wing disc cells respond to injury during regeneration processes, they would be able to compare the identity of their cells of interest with the wild-type imaginal disc cells in our database.
I hope that our database can provide a good reference point for the community
When doing the research, did you have any particular result or eureka moment that has stuck with you?
MD After I assigned the disc cells correctly to the pouch/hinge region, I was very happy to see that fine patterning processes are well represented in our single cell data. This gave me a sense of connection and also deep respect for the classical works on pattern formation, which I had previously only learned from textbooks.
And what about the flipside: any moments of frustration or despair?
MD I started as the only student working on computation in our lab and needed to learn everything from zero. My lab mates are all excellent experimentalists but they cannot help me with computational problems. It took a while to grow out of the loneliness but I have become more confident now.
So what next for you after this paper?
MD I have just finished my second year as a postgraduate student. I am now pursuing a few quantitative biology projects for which we already have data and a priority for me is to further sharpen my computational and mathematical skills. Hopefully, I can share new exciting stories in a few years when I graduate with my PhD.
Where will this work take the Yan lab?
YY The scrib mutant cells are very interesting, because when they are generated as mosaic clones in the wing discs they behave very differently and undergo cell death through a cell competition process. Building upon this work, we are now trying to better understand the scrib mutant clonal cells, and how different signalling activities contribute to their cell plasticity at the single cell level and eventually alter their growth outcome.
Finally, let’s move outside the lab – what do you like to do in your spare time in Hong Kong?
YY: I have a 4-year-old son and I am expecting another baby in December, so my activities outside the lab revolve around parenting. I find the parenting experience extremely helpful in that I am much more patient with students now than before.
MD: Hong Kong is a surprisingly great place for outdoor activities like hiking and sailing, which I like. I also like to play soccer and computer games.
 (No Ratings Yet)
(No Ratings Yet) Cell adhesion molecules (CAMs) of the plasma membrane physically tie cells together into tissues, both via interaction of cells with other cells or with ECM (extracellular matrix); they also constitute hubs of information exchange with neighbouring cells and the environment. CAMs mediate a broad range of biological functions and their aberration contributes to a range of diseases, such as cancer progression or aberrant growth/regeneration of nerves.
Cell adhesion molecules (CAMs) of the plasma membrane physically tie cells together into tissues, both via interaction of cells with other cells or with ECM (extracellular matrix); they also constitute hubs of information exchange with neighbouring cells and the environment. CAMs mediate a broad range of biological functions and their aberration contributes to a range of diseases, such as cancer progression or aberrant growth/regeneration of nerves.
 (1 votes)
(1 votes)