Join invited speakers and showcase your work to celebrate the 30th HGM. We particularly welcome presentations from budding scientists to foster networking and collaboration opportunities.
This 2-day meeting will end with the Peter Thorogood Memorial Lecture by Prof. William Harris.
29-30 January, 2018 – UCL Great Ormond Street Institute of Child Health, London, UK
Abstract submission for oral or poster presentation: 15th December, 2017
Conference Registration: 8th January, 2018
BackgroundtotheMeeting
Following the 1987 BSDB Meeting on “Craniofacial Development” organized by Peter Thorogood and Cheryll Tickle, the organizers and other colleagues (Mark Ferguson, Andrew Lumsden, Gudrun Moore, Alisdair Ivens, Julian Lewis and Gillian Morriss-Kay) decided to start holding annual informal meetings to increase communication between laboratories working on various aspects of the development of the head, to offer the opportunity to young scientists to present their work in a friendly and stimulating environment and promote collaborative research. The group has met annually since 1989, and the initial number of participants, about 20 people, rose very rapidly through word of mouth with more groups asking to be included in the mailing list. Over the last few years there have always been over one hundred people attending the meeting.
A postdoctoral research position is available starting in 2018 with Asst. Prof. Timothy Saunders’ group at the Mechanobiology Institute, Singapore. The Saunders lab has been active since 2013 and studies the fundamental processes shaping organs and tissues during development.
One major area of study in the lab is spatial position – how do developing embryos know where to position boundaries? One mechanism is the morphogen gradient, but, despite significant work over the past 20 years, there is still little reliable data about the dynamics of morphogen gradient formation. We are part of a major five-year grant focused on understanding the dynamics of morphogen gradient formation in Zebrafish embryogenesis. The project is a close collaboration between experimentalists, biophysicists and computational biologists. We are looking for a talented post-doctoral researcher interested in working at the interface of developmental biology and biophysics. In particular, the position will involve extensive imaging and image analysis of the early Zebrafish embryo, including light-sheet microscopy. This offers an exciting opportunity for a dedicated researcher to be part of a genuinely interdisciplinary project that will advance our fundamental understanding of development.
Candidates should have strong experience in at least one of the following, and display a willingness to learn the other: (1) advanced microscopy techniques and imaging developing organisms; and/or (2) experience with image analysis and handling large datasets.
The Saunders’ lab is a young group at the Mechanobiology Institute. This provides an exciting prospect for a motivated post-doc to be involved in developing research directions. The post-doc will also be expected to guide and help the graduate students in the laboratory. More information can be found at: http://labs.mbi.nus.edu.sg/mod/.
Qualifications:
1) A Ph.D in Biophysics, Developmental Biology, Computational Biology, or related subject.
2) At least one first-author paper in English submitted to an international peer-reviewed journal.
3) Experience in: (i) Advanced microscopy; (ii) Image analysis and handling large datasets; and (iii) Imaging developing organisms.
Salary and benefits are commensurable to educational qualifications and working experience of the candidates. Benefits include annual leave, medical and flexi-benefits, etc.
Please submit your curriculum vitae and a statement about your research interests to
Lee lab has been studying on cell specification process using human pluripotent stem cells, particularly nervous system and skeletal muscle cells to study human diseases and to develop new drugs.
We are looking for highly motivated postdoc(s) who have experience on one of the following fields, including Alzheimer’s disease and tau pathology.
Previous experience on human pluripotent stem cells is not required (but basic cell culture experience will be great), and we value more on the ‘non-stem cell’ expertise. Please send your application (CV and three reference contact info) to Gabsang Lee (leelabjob@gmail.com).
Neurodevelopmental disorders are a group of different conditions in which the development of the central nervous system is disturbed. This includes developmental brain dysfunction, which can manifest as neuropsychiatric problems (Autism Spectrum Disorders, schizophrenia, fragile-X syndrome, down syndrome), or impaired motor function, learning, language or non-verbal communication. The generation of the appropriate diversity of neural cell types, their migration to correct sites in the brain and the establishment of precise connectivity with target cells are key developmental processes that may go awry, leading to brain dysfunction and neurological diseases.
During this advanced 3-week course, we will provide participants with a comprehensive theoretical and experimental/practical training on the most advanced methodologies and models in developmental neurobiology, in health and disease. World experts (both seniors and juniors) in the field of neural stem cells, brain patterning, neuronal migration, axon guidance or the genetics of human neurodevelopmental disorders will give stimulating lectures and design mini-projects to be carried out by the students.
This will give students a unique opportunity to become familiar with cutting-edge in vivo gene manipulation methods (electroporation, Crispr/Cas9, genetics…), 3D imaging technologies (SPIM and light sheet microscopy, tissue clearing, two-photon imaging…), time-lapse microscopy and cell tracking, advanced pluripotent stem cell culture (iPSCs, cortical progenitors and neurons derived thereof, organoids, organotypics…).
Deadline: 18 December 2017 (Midnight Brussels time)
The Johnson Lab in the Department of Developmental Biology at Washington University School of Medicine (http://devbio.wustl.edu) is seeking applications for NIH funded Research Scientist positions. Our lab uses multiple model systems to uncover molecular mechanisms of muscle development and disease. We are looking for highly motivated individuals to begin new projects using mouse models and human ES cells to probe mechanisms of muscle disease.
DUTIES:
Assists in the training and development of technicians, acting as a group leader.
Independently generates mouse strains and collects and analyses histology data.
Develops human ES cell differentiation methods using published protocols.
Performs complex analysis/projects according to research protocols, explaining methods and procedures to other technicians.
QUALIFICATIONS:
Master’s degree with 2 years of experience in a laboratory setting (or equivalent combination of education and experience equaling 7 years)
Experience with mouse husbandry and mammalian cell culture
Please send a cover letter, CV, and list of 3 references to anjohnson@wustl.edu
Glaucoma is a devastating retinal degenerative disease without effective treatments, and its causes remain elusive. Our laboratory uses mice as a model to study glaucoma pathogenesis at the molecular level. A NIH-funded postdoctoral position is available immediately to investigate glaucoma pathogenesis in mice. In addition, our laboratory also studies the molecular mechanisms underlying adult stem cell self-renewal and differentiation. The candidate also has a unique opportunity to investigate the potential of using stem cells to treat retinal degenerative diseases. Our research has been published in high-profile journals, including Science, Nature, Cell Stem Cell and PNAS. Preferably, the candidate has a Ph.D. degree in molecular biology, developmental biology and/or genetics (research experience with mouse research and vision research is a plus). We offer highly competitive salary plus medical benefits. Our lab is accessible to state-of-art expert-run facilities, including high throughput sequencing, microarrays, proteomics, histology, FACS and imaging. For more information about the research in Dr. Xie’s laboratory, please visit the website: http://www.stowers.org/faculty/xie-lab. If interested and any questions, please send an email to Dr. Ting Xie attgx@stowers.org.
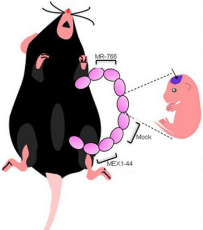
Zika infection in humans is associated with birth defects including microcephaly. Zika has two major lineages – the Asian lineage, which has been associated with birth defects, and the African lineage, which has not – but the relative effects of each strain on brain development, and the effects of the related dengue virus that co-circulates with Zika, are not understood. This week we feature a paper published in the latest issue of Development that uses a mouse model to compare the individual effects of these viruses. Co-first authors, Qiang Shao and Stephanie Herrlinger, and PI Jian-Fu (Jeff) Chen told us more.
Qiang Shao, Stephanie Herrlinger and Jeff Chen
Jeff, can you give us your scientific biography and the main questions your lab is trying to answer?
JFC My lab is interested in mechanisms underlying microcephaly and neurodegeneration. We use mouse and human pluripotent stem cell-derived neural cells to model neurological disorders followed by mechanistic studies at molecular, cellular, and circuit levels.
Qiang and Stephanie, how did you come to join the Chen lab?
SH I have always been very passionate about studying the brain and I was looking for a mentor who was as enthusiastic as I was. Jeff’s previous work studying brain development and neural tube defects gave me the opportunity to explore brain development and embryology, and his enthusiasm for understanding biology is infectious. After the first week of my rotation with his lab, it was obvious that I was going to do my PhD with Jeff.
QS I did my PhD in China, focusing on the discovery of antiviral genes and drugs. I am always passionate about brain development and neuro-infectious disease. Before my graduation, one of my committee members, Dr. He, recommended me to Jeff for a postdoc position, knowing that he is an expert in neurodevelopmental disorders. After a few rounds of interviews, Jeff offered me this position. And here I am studying Zika virus and microcephaly.
What was known about the differences between the two lineages, both in the lab and in the human population, before your work?
JFC We knew that Asia Zika virus (ZIKV) is sufficient to cause microcephaly and additional brain abnormalities from both animal models and the human population. On the other hand, Africa ZIKV had not yet been associated with any birth or brain defects. Before our work, we also knew there was about a 110 amino acid difference between the Africa ZIKV isolate and Asia isolate in our lab, but we had no idea about their comparative toxicity and their impact to the developing brain.
Can you give is the key results of the paper in a paragraph?
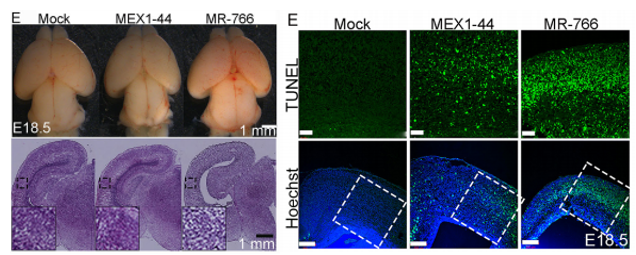
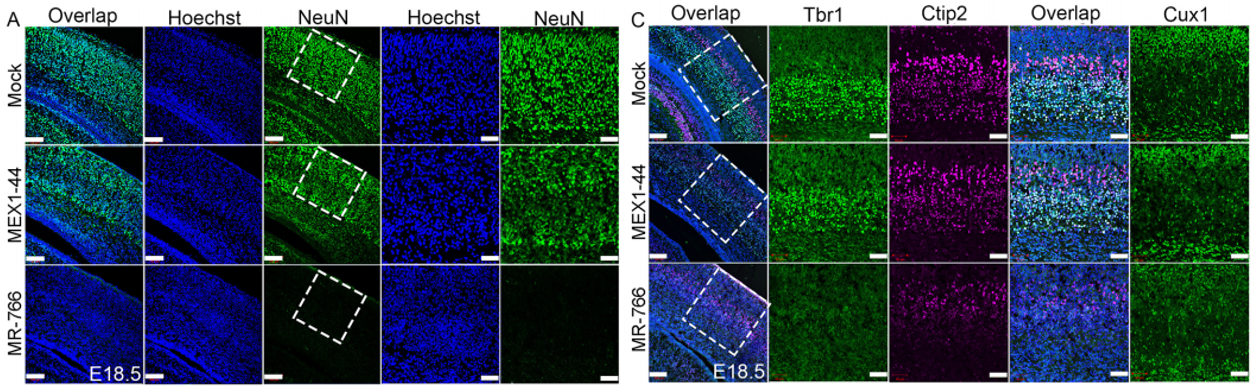
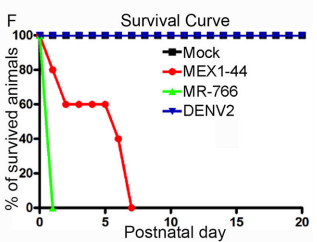
JFC The key findings are that Africa Zika virus (ZIKV) grows faster, causes more cell death in neural progenitor cells (NPCs) and neurons, and leads to more severe brain damage and postnatal death than Asia ZIKV. Both viruses similarly infect NPCs and trigger microglial activation and astrogliosis. Meanwhile, Dengue virus also infects NPCs and grows robustly in the developing brain, but fails to cause brain damage and postnatal death. Therefore, Africa ZIKV and Asia ZIKV have some intrinsic differences that account for their differential impacts on the developing brain.
Africa Zika causes a smaller cortex and increased progenitor and neuronal death than Asian Zika. From Figures 4 and 7, Shao et al, 2017.
Why did you test this hypothesis in a mouse model as opposed to using human cerebral organoids?
JFCWe previously have established the first postnatal microcephaly mouse model associated with ZIKV infection (Shao et al., Development, 2016). It is handy for us to use this system. It is absolutely important to further test this hypothesis using human cerebral organoids, which have the advantage of modelling early human brain development.
SH Cerebral organoids are a great system to study NPCs and NPC behaviours at early stages; however, the developing brain includes a diverse set of neural progenitor, immune, vascular, and glial cell types. With our mouse model, we are able to specifically ask how the heterogeneous cell populations of the brain responds and contributes to ZIKV infection pathology and disease progression.
Africa Zika causes more neuronal loss than Asia Zika. From Figure 5, Shao et al, 2017.
You mention cases of co-infection between Zika and dengue, do you have plans to model this in mice and what effect do you hypothesise it will have on neural development?
JFC We would like to model it and we expect that co-infection between Zika and dengue will yield an intermediate brain defect.
What do you hypothesise the genetic/molecular basis might be for the different in virulence between the strains?
JFC We expect that certain mutations in one or more structural/non-structural viral proteins accounts for the difference. We are actively pursuing this direction now.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
SH When we first saw the much more dramatic pathology of the African ZIKV strain on the developing brain, we were very surprised! The fact that this virus was not previously associated with birth defects was a big part of why we hypothesized that the Asian-ZIKV would be more deleterious than the African-ZIKV strain.
QS It was how the ZIKV African strain caused embryonic lethality that struck me the most. Before this study, there was no evidence suggesting an association between ZIKV African strain and microcephaly. I was expecting to see a mild phenotype in ZIKV African strain infected mouse embryos compared to Asian strain. It turned out that none of the ZIKV African strain infected embryo survived to postnatal stage. I was shocked by the phenotype of embryonic lethality. After this project, I realised unexpected results often lead to interesting and important discovery.
Postnatal survival of mice infected with the different viruses. From Figure 4, Shao et al, 2017.
And what about the flipside: any moments of frustration or despair?
SH It was frustrating at first to find that the African-ZIKV infected embryos did not survive gestation as the Asian-ZIKV infected embryos did. We were initially concerned that it was due to some complication of the surgeries as they can be arduous and the embryos are sensitive.
QS ZIKV African and Asian strain side by side comparison surgery is the most difficult part of this study. Statistical analysis requires three biological replicates. So I had to inject three embryos with mock, ZIKV-African, and ZIKV-Asian, respectively, which means nine embryos in one litter. This presents with many technical and experimental challenges including the potential of early embryo loss. It took me a whole month to optimize surgical conditions such as polishing glass needles, reducing viral titer and adjusting injection sites. Fortunately, through a long process of trial and error, I was able to obtain enough samples to work with.
Infection of embryonic mice. From Figure 4, Shao et al, 2017.
What are your career plans following this work?
SH I successfully received a predoctoral to postdoctoral transition award from the NIH (D-SPAN F99/K00) to study the impact that ZIKV has on post-transcriptional regulation in infected neural progenitor cells. In the coming year I plan to graduate and move on to my postdoctoral career.
QS I am currently writing a research proposal to further study the mechanism underlying ZIKV caused neurovascular defect. Specifically, I am interested in understanding the role of individual ZIKV protein in neurovascular development disruption. This work will provide important molecular mechanism of ZIKV pathogenesis.
And what next for the Chen lab?
JFC We are trying to figure out the molecular mechanisms underlying Zika virus-induced microcephaly and associated neurological disorders.
Finally, what do you three like to do when you are not in the lab?
JFC I always like to spend more time with my family.
SHI like to spend time going to see live music, my family, hiking, and travelling.
QS I enjoy spending time with my partner and friends. I love cooking at home. It’s kind of like doing experiments in the kitchen with different ingredients. I find those moments very relaxing and full of delight.
The Mokalled lab in the Department of Developmental Biology at Washington University School of Medicine is hiring at all levels (http://www.mokalledlab.com/). Our lab uses zebrafish and mouse model systems to study neural regeneration after spinal cord injury or disease. Candidates with enthusiasm for neuroscience, regenerative biology, and zebrafish research are encouraged to forward a cover letter, CV, and list of 3 or more references to mmokalled@wustl.edu.
We are recruiting 1-2 PhD students (to begin Fall 2018) and a postdoctoral fellow (position immediately available) to study cell shape changes and rearrangement and underlying cytoskeletal regulation during organ formation in Dr. SeYeon Chung’s newly established lab at Louisiana State University.
We use the genetically tractable Drosophila salivary gland as a model system to understand how a flat sheet of epithelial cells becomes a three-dimensional tubular structure during development. The project will employ a combination of fly genetics, advanced imaging and image analysis, and molecular and biochemical approaches.
Related publication: Chung, S., et al., (2017) Uncoupling apical constriction from tissue invagination. eLife 6:e22235.
Grad students: Highly motivated candidates with a strong undergraduate degree in any area related to the biological sciences are encouraged to apply.
Postdocs: Highly motivated candidates who recently obtained or are about to obtain a PhD in a field of cell biology, developmental biology or biochemistry are encouraged to apply.
If interested, please send your CV, a brief description of your research interests, and contact information of three references to Dr. SeYeon Chung (seyeonchung@lsu.edu).
Here are the highlights form the current issue of Development:
Cross-border control of stem cell behaviour
Cell identity and proliferation differs between organs, raising the question of how cells at interorgan boundaries are regulated to maintain organ integrity. On p. 4091, Don Fox and colleagues identify a specialised transition zone at the midgut/hindgut boundary in the Drosophila intestine. This ‘hybrid zone’, which shows gene expression profiles from both organs, changes in size during development but is maintained into adult life, and cells within it contribute to both midgut and hindgut tissue. The authors describe a new population of stem cells – the organ-boundary intestinal stem cells (OB-ISCs) – that reside in the midgut immediately adjacent to the hybrid zone and show slower division rates. Injury to the hybrid zone increases proliferation of these OB-ISCs, and if the injury is severe enough, hyperplastic OB-ISCs can cross the boundary and invade the hindgut. The authors find that OB-ISC proliferation is induced by release of the JAK-STAT ligand Unpaired-3 from the hindgut and the hybrid zone following injury. The hybrid zone therefore serves as a focal point of interorgan interaction to influence the behaviour of OB-ISCs and preserve the midgut/hindgut distinction, raising the possibility that interorgan regulation of stem cell behaviour may be a common mechanism to maintain organ identity.
Migrating interneurons get active
During the development of the vertebrate central nervous system, interneuron precursors migrate extensively before they reach their final destination in the brain, where they then differentiate and become integrated into functional circuits. It is known that developing interneurons are sensitive to neurotransmitters but now, on p. 4125, Ronald Jabs and co-workers show that interneuron precursors require synaptic input for correct migration. Focussing on a population of cerebellar molecular layer interneurons in mice, the authors use time-lapse imaging and patch-clamp recordings to determine the morphological and electrophysiological characteristics of migrating interneuron precursors. Their analyses reveal that interneuron precursors exhibit spontaneous postsynaptic currents and receive both glutamatergic and GABAergic input as they migrate. Ultrastructural studies further demonstrate the presence of synaptic structures on these cells, and the authors also show that the density of synaptic elements increases along the migratory route of these interneuron precursors. Finally, the researchers report that blocking synaptic transmission, using tetanus toxin and Co2+ to abrogate presynaptic release, perturbs migration; both the speed and directionality of migration are reduced. Together, these findings reveal that the direct synaptic innervation of migrating interneuron precursors regulates their migratory behaviour and highlights an important and unprecedented role for synapses during neuronal pathfinding.
Zika: strain-specific impacts on brain development
Infection with Zika virus during pregnancy can lead to severe birth defects in humans, including microcephaly. Zika has two major lineages – the Asian lineage, which has been associated with birth defects, and the African lineage, which has not – but the relative effects of each strain on brain development, and the effects of the related dengue virus that co-circulates with Zika, have not been addressed. On p. 4114, Jian-Fu Chen and colleagues address this problem by performing intracerebral inoculation with Zika and dengue virus on embryonic mouse brains and comparing their effects on neural development. They show that both dengue and Zika viruses cause microcephaly through impaired neural progenitor proliferation and increased neuronal apoptosis, though the effect is much greater for Zika than dengue. Surprisingly, given the apparent absence of virus-related pathology in affected human populations, the African strain grows faster and causes greater progenitor and neuronal cell death, and higher postnatal mortality, than the Asian lineage. This study generates insights into the neurodevelopmental phenotypes generated by these viruses, and provides a foundation for future investigations into the molecular and genetic causes of Zika pathogenesis.
PLUS:
Fibroblast growth factors: key players in regeneration and tissue repair
Both regeneration and repair are orchestrated by a highly coordinated interplay of different growth factors and cytokines. Among the key players are the fibroblast growth factors (FGFs), which control the migration, proliferation, differentiation and survival of different cell types. In addition, FGFs influence the expression of other factors involved in the regenerative response. In their Review, Sabine Werner and colleagues summarize current knowledge on the roles of endogenous FGFs in regeneration and repair in different organisms and in different tissues and organs.
The evolution of cortical development: the synapsid-diapsid divergence
During evolution, the cortex appeared in stem amniotes and evolved divergently in two main branches of the phylogenetic tree: the synapsids (which led to present day mammals) and the diapsids (reptiles and birds). Comparative studies in organisms that belong to those two branches have identified some common principles of cortical development and organization that are possibly inherited from stem amniotes and regulated by similar molecular mechanisms. These comparisons have also highlighted certain essential features of mammalian cortices that are absent or different in diapsids and that probably evolved after the synapsid-diapsid divergence. In his Review, Andre Goffinet discusses this data and provides an evolutionary perspective on cortical neurogenesis, neuronal migration and cortical layer formation and folding.
Developing a sense of touch
The sensation of touch is mediated by mechanosensory neurons that are embedded in skin and relay signals from the periphery to the central nervous system. During embryogenesis, axons elongate from these neurons to make contact with the developing skin. Concurrently, the epithelium of skin transforms from a homogeneous tissue into a heterogeneous organ that is made up of distinct layers and microdomains. Throughout this process, each neuronal terminal must form connections with an appropriate skin region to serve its function. In their Review, Blair Jenkins and Ellen Lumpkin present current knowledge of the development of the sensory microdomains in mammalian skin and the mechanosensory neurons that innervate them.
 (1 votes)
(1 votes) (No Ratings Yet)
(No Ratings Yet)


 Cell identity and proliferation differs between organs, raising the question of how cells at interorgan boundaries are regulated to maintain organ integrity. On p.
Cell identity and proliferation differs between organs, raising the question of how cells at interorgan boundaries are regulated to maintain organ integrity. On p.  During the development of the vertebrate central nervous system, interneuron precursors migrate extensively before they reach their final destination in the brain, where they then differentiate and become integrated into functional circuits. It is known that developing interneurons are sensitive to neurotransmitters but now, on p.
During the development of the vertebrate central nervous system, interneuron precursors migrate extensively before they reach their final destination in the brain, where they then differentiate and become integrated into functional circuits. It is known that developing interneurons are sensitive to neurotransmitters but now, on p.  Infection with Zika virus during pregnancy can lead to severe birth defects in humans, including microcephaly. Zika has two major lineages – the Asian lineage, which has been associated with birth defects, and the African lineage, which has not – but the relative effects of each strain on brain development, and the effects of the related dengue virus that co-circulates with Zika, have not been addressed. On p.
Infection with Zika virus during pregnancy can lead to severe birth defects in humans, including microcephaly. Zika has two major lineages – the Asian lineage, which has been associated with birth defects, and the African lineage, which has not – but the relative effects of each strain on brain development, and the effects of the related dengue virus that co-circulates with Zika, have not been addressed. On p.  Both regeneration and repair are orchestrated by a highly coordinated interplay of different growth factors and cytokines. Among the key players are the fibroblast growth factors (FGFs), which control the migration, proliferation, differentiation and survival of different cell types. In addition, FGFs influence the expression of other factors involved in the regenerative response. In their
Both regeneration and repair are orchestrated by a highly coordinated interplay of different growth factors and cytokines. Among the key players are the fibroblast growth factors (FGFs), which control the migration, proliferation, differentiation and survival of different cell types. In addition, FGFs influence the expression of other factors involved in the regenerative response. In their  During evolution, the cortex appeared in stem amniotes and evolved divergently in two main branches of the phylogenetic tree: the synapsids (which led to present day mammals) and the diapsids (reptiles and birds). Comparative studies in organisms that belong to those two branches have identified some common principles of cortical development and organization that are possibly inherited from stem amniotes and regulated by similar molecular mechanisms. These comparisons have also highlighted certain essential features of mammalian cortices that are absent or different in diapsids and that probably evolved after the synapsid-diapsid divergence. In his
During evolution, the cortex appeared in stem amniotes and evolved divergently in two main branches of the phylogenetic tree: the synapsids (which led to present day mammals) and the diapsids (reptiles and birds). Comparative studies in organisms that belong to those two branches have identified some common principles of cortical development and organization that are possibly inherited from stem amniotes and regulated by similar molecular mechanisms. These comparisons have also highlighted certain essential features of mammalian cortices that are absent or different in diapsids and that probably evolved after the synapsid-diapsid divergence. In his  The sensation of touch is mediated by mechanosensory neurons that are embedded in skin and relay signals from the periphery to the central nervous system. During embryogenesis, axons elongate from these neurons to make contact with the developing skin. Concurrently, the epithelium of skin transforms from a homogeneous tissue into a heterogeneous organ that is made up of distinct layers and microdomains. Throughout this process, each neuronal terminal must form connections with an appropriate skin region to serve its function. In their
The sensation of touch is mediated by mechanosensory neurons that are embedded in skin and relay signals from the periphery to the central nervous system. During embryogenesis, axons elongate from these neurons to make contact with the developing skin. Concurrently, the epithelium of skin transforms from a homogeneous tissue into a heterogeneous organ that is made up of distinct layers and microdomains. Throughout this process, each neuronal terminal must form connections with an appropriate skin region to serve its function. In their