Bill Harris, winner of the BSDB Waddington Medal 2017
Posted by BSDB, on 13 April 2017

 Bill is Canadian, but underwent his scientific education and early career in the U.S., where he did his B.A. in Biophysics (University of California, Berkeley; 1972), his Ph.D. on “Color vision in Drosophila” in the group of Seymour Benzer at the California Institute of Technology (Pasadena; 1972-76), carried out his postdoctoral research in the laboratory of David Hubel and Torsten Wiesel at the Dept. of Neurobiology, Harvard Medical School (1976-80), and joined the faculty of the Dept. of Biology, University of California (San Diego; 1980). He remained in San Diego until 1997, when he moved to the UK to take on a position as Professor of Anatomy at the University of Cambridge and, since 1999, Head of the Department of Anatomy (which became the Department of Physiology, Development and Neuroscience in 2006).
Bill is Canadian, but underwent his scientific education and early career in the U.S., where he did his B.A. in Biophysics (University of California, Berkeley; 1972), his Ph.D. on “Color vision in Drosophila” in the group of Seymour Benzer at the California Institute of Technology (Pasadena; 1972-76), carried out his postdoctoral research in the laboratory of David Hubel and Torsten Wiesel at the Dept. of Neurobiology, Harvard Medical School (1976-80), and joined the faculty of the Dept. of Biology, University of California (San Diego; 1980). He remained in San Diego until 1997, when he moved to the UK to take on a position as Professor of Anatomy at the University of Cambridge and, since 1999, Head of the Department of Anatomy (which became the Department of Physiology, Development and Neuroscience in 2006).
Bills achievements are best summarised in the nomination letter put forward by Sarah Bray, Michael Bate, Nancy Papalopulu, Daniel St Johnston and Steve Wilson:
“Through his passion for science, his leadership and his mentoring of people at all career stages, Bill has made outstanding contributions to Developmental Biology. Working at the interface of Developmental Biology and Neuroscience, he has championed the field within Cambridge, across the UK and throughout the world. His deep interest and scientific enthusiasm have led to major insights in the field of neuronal specification and wiring.
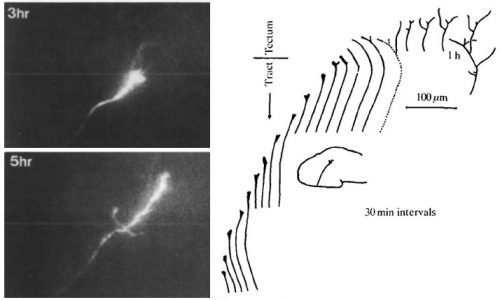
Bill has made many important contributions to our understanding of visual system development, which has been his focus throughout his career. Time and again, he has pioneered new fields of research. His early discoveries in the USA helped establish basic principles underlying axon guidance in brain wiring and revealed key mechanisms regulating cell differentiation in the retina. Starting with the identification in Drosophila of the sevenless gene (with Seymour Benzer), he went on to discover ways that axons in the brain are guided to the vicinity of their appropriate targets in the absence of neural activity, being one of the first to suggest there might be local chemotactic cues that guide retinal axons from a distance (see Figure 1; Dingwell et al., 2000, J Neurobiol 44, 246ff. – LINK). He used the developing Xenopus visual system to find the first vertebrate homologues of genes that influenced fate choice in Drosophila, including Notch, ASH, and ATH, leading to a new and fruitful research direction for Developmental Biologists.
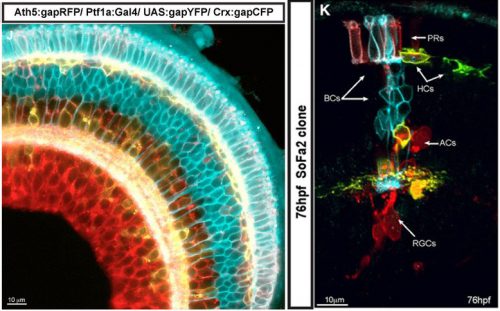
After moving to Cambridge in 1997, he built on these earlier observations. He established the retinal ciliary marginal zone as a powerful model to study not only retinal development per se, but also mechanisms controlling stem cells and the pathways regulating differentiation. Perpetually self-renewing and proliferative, this neuroepithelium at the perimeter of the retina in amphibians and fish gives rise to cells that are spatially ordered with respect to cellular development. Combining in vivo lipofection strategies in Xenopus retina with genetic approaches in zebrafish, Bill has uncovered roles for the cell cycle, metabolism and stochasticity in fate determination. A pioneer in the field of live imaging in developing systems, he made the very first time-lapse movies of axons growing in the brain. He has harnessed emerging technology to distinguish between different hypotheses. For example, by watching lineages evolve in vivo he ruled out the idea that their variance was due to random cell death. Through such cutting-edge studies he has shown how cell divisions, cell lineages and cell polarizations contribute to the process of retinal neuron specification (Fig. 2; Agathocleous & Harris, 2009, Annu Rev Cell Dev Biol 25, 45ff. – LINK). In doing so, he championed the establishment of the Cambridge Advanced Imaging Centre, whose main focus is on techniques that enable gentle deep imaging of cells in developing animals, and has forged strong links with physicists to develop powerful models.
Bill has made important contributions to the community. He is an enormous presence in Cambridge, heading one of the major biology departments and taking a lead in the Cambridge Neuroscience initiative. Under his leadership and long-term vision, the Departments of Anatomy and Physiology were merged, establishing “PDN” where the letter D stands for Development. Bill is on the Advisory committee for the Gurdon Institute, and has also been instrumental in the establishment and renewal of the Wellcome Trust 4-year PhD Programme in Developmental Biology. As well as training first-rate researchers in his own lab (23 of whom have PI positions around the world), he has, as Department Head, nurtured many developmental biologists at different stages in their career, housing early researchers (e.g. Fiona Wardle), recruiting talented lecturers (e.g. Clare Baker, Benedicte Sanson, Kristian Franze) and supporting established leaders (e.g. Andrea Brand, Magda Zernicka-Goetz). Despite the burdens of being Department Head, Bill has retained a major teaching role throughout, with lectures and practicals introducing fundamentals of neural development to undergraduates at all stages. As organizer of a number of international conferences, he has included significant themes in Developmental Biology. He is also editor in chief of “Neural Development” and on the editorial boards for “PLoS Biology”, “Cell” and “Molecular Neuroscience”. Among his many other talents, Bill is an artist who also communicates his scientific vision through his paintings, as illustrated by his impression of a young zebrafish retina (see Figure 3).”

To add to this, the high quality of Bill’s work is reflected in his many honours, fellowships and awards which include being a Fellow of the Academy of Medical Sciences (2007), Fellow of the Royal Society (2007) and Member of EMBO (2012). With respect to translational biology, he was founder of the drug discovery company DanioLabs (2002), which was successfully sold to VASTox (now Summit) plc, a leading UK biotechnology company [LINK]. Furthermore, for many of us, Bill has become an inspiring teacher, especially through his textbook Development of the Nervous System (Sanes et al., 3rd edition, 2011, Academic Press – LINK) which was a true eye opener: a highly entertaining read laying essential foundations for conceptual thought about the field and its many facets and directions.

Finally, Bill’s passion for ice hockey deserves mentioning (Fig. 4). He decided to share it with the Cambridge community by founding the Cambridge Leisure and Ice Centre as a community-led initiative in 2001, which he still chairs today. The Cambridge Ice Arena will be the key result of the initiative and is scheduled to open late 2017.
The BSDB would like to congratulate Bill for his life achievements and for being the well-deserved awardee of the 2017 Waddington Medal. The Waddington Medal lecture is available on the BSDB YouTube channel and website, and an interview with Bill was published in Development.
 (6 votes)
(6 votes)

 (No Ratings Yet)
(No Ratings Yet)
 His Beddington Medal talk described the outcome of his most successful PhD project. About the background Erik explained: “The Drosophila segmentation cascade is a paradigmatic example of a developmental gene regulatory network, used to gain insight into transcriptional regulation in all animals… Using spatial information from graded domains of “gap” gene expression, seven “pair-rule” genes are expressed in periodic patterns of seven stripes each .. [which] then work in combination to specify precisely-phased 14 stripe patterns of “segment polarity” genes. These output patterns form the template for the segmental organisation of the insect body…[Although] the pair-rule genes have been studied in Drosophila for over 30 years,… there is still no good systems-level understanding of their regulatory interactions“.
His Beddington Medal talk described the outcome of his most successful PhD project. About the background Erik explained: “The Drosophila segmentation cascade is a paradigmatic example of a developmental gene regulatory network, used to gain insight into transcriptional regulation in all animals… Using spatial information from graded domains of “gap” gene expression, seven “pair-rule” genes are expressed in periodic patterns of seven stripes each .. [which] then work in combination to specify precisely-phased 14 stripe patterns of “segment polarity” genes. These output patterns form the template for the segmental organisation of the insect body…[Although] the pair-rule genes have been studied in Drosophila for over 30 years,… there is still no good systems-level understanding of their regulatory interactions“.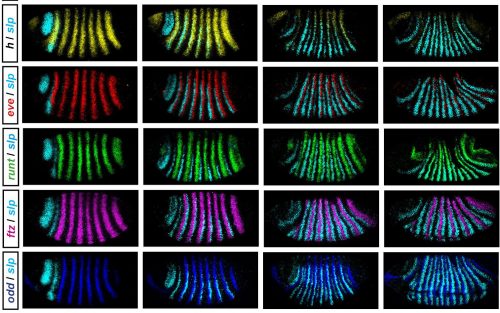




 Retinal degenerative diseases are the leading causes of blindness worldwide. In their
Retinal degenerative diseases are the leading causes of blindness worldwide. In their  Prostate organogenesis is a complex process that is regulated by androgens and subsequent mesenchyme-epithelial interactions. In their
Prostate organogenesis is a complex process that is regulated by androgens and subsequent mesenchyme-epithelial interactions. In their  As every year, the
As every year, the