Here is some developmental biology related content from other journals published by The Company of Biologists.
Xenopus as a developmental model of neuroblastoma
Neuroblastoma (NB) is a paediatric form of cancer derived from the sympathetic nervous system. Recent genome-wide sequencing data suggest that often NB does not have a clear genetic cause, leading the authors to hypothesize that NB results from aberrations of normal development. To test this hypothesis, Anna Philpott’s group used a population of anteroventral noradrenergic (AVNA) cells from Xenopus embryos. These cells share several features with mammalian sympathetic neurons, including the expression of noradrenergic-associated genetic markers such as the achaete-scute complex-like 1 (Ascl1) gene, which encodes a transcriptional driver of neurogenesis. By comparing AVNA and NB cells, the authors found that, whereas Ascl1 is only transiently expressed in AVNA cells, it is aberrantly maintained in NB, where it is phosphorylated on multiple serine-proline sites. The authors then show that differentiation of AVNA cells is enhanced by dephosphorylated Ascl1. Moreover, this process is inhibited by experimental manipulations of NB-associated genes, but, interestingly, dephosphorylation of Ascl1 is able to overcome this inhibition. This work demonstrates that Xenopus AVNA cells represent a unique system to study sympathetic nervous system development and its relationship to NB. Moreover, it suggests that Asc11 phosphorylation might promote stalled differentiation leading to NB, thus identifying a potential target for therapeutic purposes. Read the paper here (Open Access) and the authors’ Node post here.
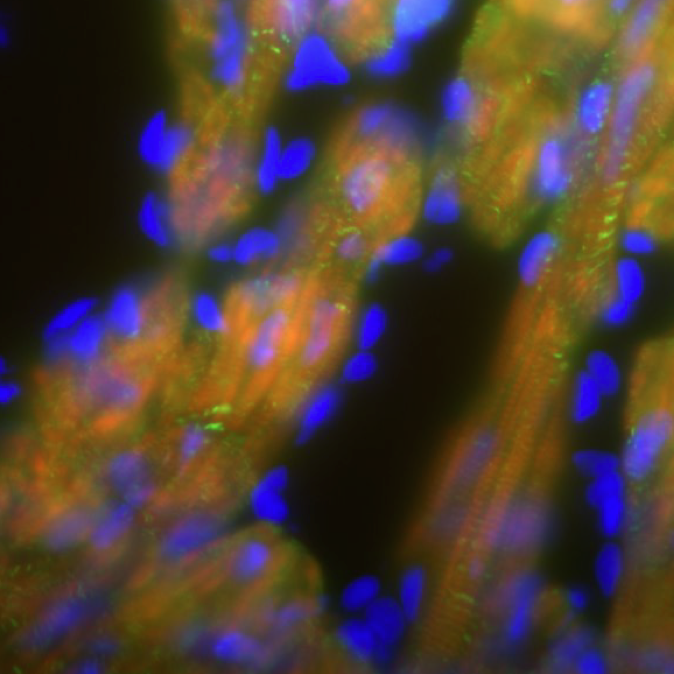
Modelling DMD-associated cardiomyopathy in patient-derived iPSCs
Duchenne muscular dystrophy (DMD) is a genetic muscular disorder characterised by progressive muscular weakness and wasting, with cardiac complications (such as dilated cardiomyopathy) and respiratory muscle failure arising in the late stage. To investigate mechanisms of dilated cardiomyopathy in DMD, Lei Yang and collaborators derived cardiomyocytes (CMs) from DMD-patient-specific induced pluripotent stem cells (iPSCs). Compared to control cells, DMD iPSC-CMs exhibited elevated cytosolic calcium, mitochondrial damage and increased cell apoptosis. To further dissect the mechanisms underlying these alterations, the authors performed transcriptional and translational analyses and identified a mitochondrially initiated molecular cascade – which involves CASPASE3 (CASP3) activation – as being responsible for the increased apoptosis in DMD iPSC-CMs. Notably, the application of the membrane sealant Poloxamer 188 could prevent calcium overload and CASP3 activation, significantly reducing apoptosis in these cells. Thus, the authors established a useful in vitro system to disclose mechanisms of cardiomyopathy in DMD and to identify molecular targets that could be pharmacologically manipulated. Read the paper here (Open Access).
Endocytosis and micropinocytosis internalise cadherin-6B in EMT
Epithelial-to-mesenchymal transition (EMT) is an integral developmental and physiological process, but can also be utilised by cancer cells at the initiation of metastasis. A requirement for EMT is the post-translational removal of adhesion proteins from the plasma membrane. Here (p. [164426]), Lisa Taneyhill and Rangarajan Padmanabhan study cadherin-6B (Cad6B) internalisation to elucidate the mechanisms of EMT in chick cranial neural crest cells. The authors found that in neural crest cells that are initiating EMT, Cad6B was detected in cytosolic puncta that were endocytic, rather than exocytic, in nature. They then identified two intracellular motifs that were potentially important for regulating Cad6B internalisation. Mutating the p120-catenin-binding (EED) motif, but not the dileucine (LI) motif, significantly increased Cad6B internalisation, supporting the idea that Cad6B is removed from the plasma membrane through endocytosis. However, although Cad6B colocalised with clathrin, the colocalisation was not exhaustive, suggesting that an additional mechanism is involved in Cad6B internalisation. Therefore, the authors used an array of pharmacological treatments to show that Cad6B was removed from the plasma membrane through both endocytosis and macropinocytosis, and that both of these processes depended on dynamin. This study demonstrates that EMT and neural crest migration require Cad6B internalisation through endocytosis and macropinocytosis. Read the paper here.
Generating an in vitro 3D cell culture model from zebrafish larvae for heart research
Spontaneously beating 3D ‘heart’ structures can be developedin vitro from larval zebrafish using a novel, fast and inexpensive method that could be employed in ecotoxicology and biomedical safety testing. Read the paper here.
My review of the Young Embryologist Network London Meeting 2015
As I write this, I am currently listening to the third very impressive senior PI talk in a row addressing ‘how to succeed in science’. This one is from an incredible woman who came through the German system, did a PhD at the EMBL in Heidelberg, did a postdoc there for about 10 minutes where she developed an incredible new model system, won loads of prizes, and is now running a large and successful lab at Cambridge basically solving every medical problem in pregnancy. In Cambridge she has started a family and been incredibly successful. She looks about 32.
So, I have decided to stop listening (and stop crying inside) and to start a review of the research talks. Hilariously, I am actually a judge of the talks this year (I have taken off marks for every time someone says ‘’so we took a system approach’’, and then speaks about a phenomenon/gene/conceptual framework that was entirely known or in place before people said this in talks. It’s not their fault of course – we are all prisoners of fashion. But here I am, I can be no other, as somebody once said). As ever, I have just picked out a few highlights. This is not because the other talks were bad, but because I have a very fat son who is 7 weeks old. So I have been getting up approximately every 2 hours for about 90 minutes every night for the last 7 weeks to feed an apparently insatiable baby who has supplanted me as the most important man in my wife’s life. I have slept through some talks. Sorry.
Vanessa Chong (Oxford)
This talk was awesome. This girl is at the end of her 2nd year (remember that). She is a student of Tatjiana Sauka-Spengler at the Wetherall Institute of Molecular Medicine. This is an institute that reverses the reprehensible modern trend, and has a translational name but does incredible basic research. She has developed a method for purifying nuclei from genetically targeted cells in zebrafish neural crest, and then conducted large-scale transcriptomics. She identified very counter-intuitive expression of neural crest developmental transcripts in non-neural crest tissues. What? But, she also found opposite strand transcription of these same genes in only the neural crest lineage. Antisense non-coding transcription is correlated with gene expression but not transcription (of neural crest genes). She is a 2nd year. Don’t know if I mentioned that. Impressive stuff.
Lizzy Ward (UCL)
A wonderfully old-fashioned bit of developmental biology examining the signalling of the notochord to the forming somites and ultimately vertebrae. She showed really nicely and elegantly that the notochord does signal to the somites, and that this signal is generic i.e. doesn’t vary in nature along the A/P axis of the notochord. It might only end up in a smaller journal (thought the last such study from the Stern lab ended up in Science), but it was beautifully presented and very elegant.
Rebecca McIntosh (KCL/UCL)
A talk on a favourite topic of mine: basal progenitors. These are proliferative cells in the CNS that unlike most CNS progenitors maintain a connection not to the apical surface, but the basal one. Such cells have expanded enormously in number in the primate neocortex and are suggested to underlie the evolution of large neocortices; they have expanded enormously in the cerebella of amniotes and are suggested to underlie the evolution of large cerebella (some wonderful work from an extremely talented and very good looking young lecturer in East London). Rebecca’s talk examined their biology in a much more simple and elegant system: the zebrafish spinal cord and hindbrain. This talk was a really nice combination of sophisticated genetic labelling and high-end microscopy, combined with really sensible and simple questions. The work quantified basal divisions in zebrafish (i.e. in a very small brain) and showed that elaborate cellular behaviours accompany their specification and patterns of division. As ever with good research, it generated more questions than it answered.
John Robert-Davis (KCL/Crick Institute)
John is a great bloke who is incredibly charismatic and depressingly bright. His thesis examined the molecular and mathematical basis of contact inhibition in Drosophila hemocytes and won the Beddington Prize for the best thesis produced in the UK last year in developmental or cellular biology. He is now a postdoc at the Crick Institute and I suspect a shoe-in for some incredible fellowship – he published his PhD work in Development and Cell. His talk was crap. That is all you need to know.
Okay, okay, it wasn’t crap. It was, in fact, utterly brilliant. And entirely predictably so. He won the prize for the best talk. And he used zorb ball (look it up) as a genuine scientific metaphor. Apparently he’s also a standup comedian. And he’s a great bloke. Urgh.
Alexander Fletcher (Oxford)
I should preface this review by saying that I am generally quite unimpressed by a lot (though certainly not all) of the modelling that is done in relation to developmental biology. I am sure that I am quite possibly completely mistaken and very very happy indeed to be corrected and saved from my own ignorance by the armies of physical scientists now being recruited into biology. Nevertheless, insert Martin Luther quote here.
I recently sat through a talk that had the sentence: ‘’of course without any inertia, you generate chaotic Turing signalling patterns.’’ And people say that recruiting physicists into developmental biology is confusing the field…
With my very poor mathematical ability (it is actually not that poor), I took the following from this other talk, as from so many others: we made a model using real data and we know how to do this (at this point the vast majority of the audience who do things like experiments become very scared – there is usually a slide with lots of maths on). We tested the model using real data. The real data supported the model. The model generated zero (this is the point of distinction between the good modelling studies and the majority) novel hypotheses. We concluded that the laws of physics apply to living things. A room full of biologists (or possibly just those who make funding decisions?) was incredibly impressed.
The final talk thus came as a wonderful surprise. It was brilliantly cynical, entertaining and I think important. It discussed how the process of writing code for models for scientific applications like developmental biology is hugely ineffective and should be assessed and rigorously examined by people who know how to write good code, before being foisted upon those who don’t. An idea whose time I hope has come. www.cs.ox.ac.uk/chaste.
Department/Location: Wellcome Trust – Medical Research Council Cambridge Stem Cell Institute, University of Cambridge
Salary: £38,511-£48,743
Reference: PS05620
Closing date: 14 June 2015
Limited funding: The funds for this post are available until 31 January 2018 in the first instance.
Three UK Research Councils; Biotechnology and Biological Sciences Research Council, Engineering and Physical Sciences Research Council and the Medical Research Council have invested £25m in research and equipment to develop the UK Regenerative Medicine Platform (UKRMP www.ukrmp.org.uk). The Pluripotent Stem Cell Platform (PSCP) is one of five UKRMP Hubs formed to tackle critical challenges in developing regenerative treatments from laboratory discoveries. PSCP brings together researchers from several institutions to develop the processes for taking pluripotent stem cells and their derivatives from laboratory research to the manufacture of safe, consistent and effective products suitable for clinical applications.
PSCP now has an exciting opportunity for a Project Manager with experience in management and administration at the interface between academia and industry.
We are looking for a proactive and motivated individual to manage the PSCP portfolio according to industry best practice; determining and delivering to agreed scope, quality, budget and deadlines. The Project Manager will manage a number of projects concurrently and will maintain accurate project plans, work schedules, issue and risk logs to enable projects to be delivered successfully. You will coordinate regular meetings at the different partner sites. You will develop summary documents and reports, will organise meetings and will lead the daily administration of the project. You will be a confident and articulate communicator who delivers information concisely and effectively. You will have the experience and interpersonal skills to build effective relationships with funders and a range of stakeholders including industry and regulators.
Experience of finance management, budgeting, and administration in a research-led environment is essential. Experience of leading, or working as part of, a management team with accountability for group and individual performance is highly desirable.
You will be educated to at least degree level and have a further qualification (MSc, PhD or MBA).
Once an offer of employment has been accepted, the successful candidate will be required to undergo a health assessment.
To apply online for this vacancy and to view further information about the role, please visit: http://www.jobs.cam.ac.uk/job/6475. This will take you to the role on the University’s Job Opportunities pages. There you will need to click on the ‘Apply online’ button and register an account with the University’s Web Recruitment System (if you have not already) and log in before completing the online application form.
The closing date for all applications is Sunday 14th June 2015.
Please upload your Curriculum Vitae (CV) and a covering letter in the Upload section of the online application to supplement your application. If you upload any additional documents which have not been requested, we will not be able to consider these as part of your application.
Interviews will be held on the afternoon of Wednesday 1st July 2015. If you have not been invited for interview by 24th June 2015, you have not been successful on this occasion.
Please quote reference PS05620 on your application and in any correspondence about this vacancy.
The University values diversity and is committed to equality of opportunity.
The University has a responsibility to ensure that all employees are eligible to live and work in the UK.
Figure 1. “Cytoplasm”, illustration by David S. Goodsell, the Scripps Research Institute.
When contemplating the illustrations by David S. Goodsell (Figure 1), the first thing that stands out is how cells are packed full with those wonderful little machines we call proteins. They move, interact and change shape to produce cellular functions, so our ability to observe where they localize and monitor their behaviour is extraordinarily important in biology. In general, immunocytochemistry is preferred for static protein detection, while the dynamics of protein localization can be highlighted by transgenic constructs including the protein of interest fused to a fluorescent tag. Importantly, whereas tagged proteins are irreplaceable tools for live imaging at high resolution, immunocytochemistry is far from dead. Both antibodies and fluorescent tags are so widespread in use that sometimes we assume that the results we obtain must reflect the endogenous condition. As a matter of fact, when comparing data from static and dynamic protein detection techniques, inconsistencies can emerge. On one hand artifacts can derive from handling of the tissue, for example by fixation and permeabilization (Schnell et al., 2012). On the other hand, direct tagging of a protein or perturbations in its expression levels can interfere with the way proteins fold, interact and therefore localize. In a notable example, Swulius and Jensen (2012) found that YFP-tagged MreB (a bacterial actin homolog) artifactually nucleates to form long helical structures. The description of a helical cytoskeleton in rod-shaped bacteria heavily influenced this field of research for over 10 years. However, the aggregates were not found in microstructural analyses of bacteria with untagged MreB, demonstrating that the filaments were a byproduct of protein tagging.
In the current issue of Development, we contributed a report investigating a potential alternative for protein detection in living organisms, which unifies the advantages of antibodies and live imaging.
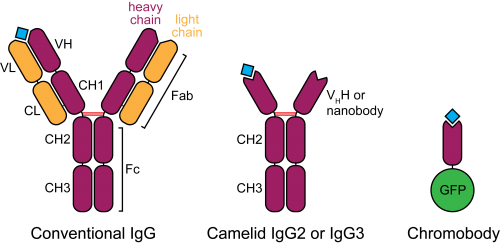
Figure 2. Schematic structure of camelid heavy-chain antibodies and chromobodies, compared to conventional IgGs.
In the late 1980s, a serendipitous discovery led to the description of heavy-chain antibodies in the plasma of camelids (Hamers-Casterman et al., 1993). Similar molecules are also found in sharks, however their advantage for animal immunity is not clear. These atypical IgGs are devoid of light chains, such that their heavy-chain variable portion (termed VHH or nanobody) can alone recapitulate antigen binding (Figure 2). This convenient property introduces the possibility of cloning entire repertoires of nanobodies and screening for binding to an antigen of interest in vitro, for example by phage display.
Nanobodies are already important reagents utilized in X-ray crystallography. They preferably bind convex surfaces, thereby helping the stabilization of protein conformations or by providing additional contact surface for protein aggregates (De Meyer et al., 2014). Their small size (15 KDa, one tenth of a conventional antibody) and high affinity are valued properties. In addition, with more and more monoclonal antibodies currently being evaluated for therapeutic application, nanobodies are becoming increasingly popular. They offer a variety of advantages, among which high tissue penetration, blood clearance, stability and solubility (Holliger and Hudson, 2005).
In a most innovative adaptation of nanobodies, Rothbauer, Zolghadr et al. (2006) fused the VHH binding moiety to a fluorescent protein and tested the resulting “chromobody” in mammalian cells. In this context, chromobodies can recapitulate the localization and original behaviour of endogenous proteins, with minimal functional interference, presumably because of their high binding turnover. Remarkably, chromobodies appear to fold correctly by establishing their only disulphide bond even in the reducing intracellular environment.
Video 1. 5 hpf zebrafish embryo injected with actin chromobody mRNA, showing localization to the plasma membrane and to the perinuclear actin cap. Centrosomes can be identified during cell division, after which the nuclear compartment and its actinic cap reassemble (arrowheads). Scale bar: 50 µm.
Inspired by these results, Ulrich Rothbauer (now at the Universität Tübingen) initiated a collaboration with our group at the Max Planck Institute for Developmental Biology, aimed at testing chromobody reagents in zebrafish. Would antigen binding be maintained in an entire organism? Would it be stable over consecutive developmental stages? We decided to focus on chromobodies binding to two proteins with well-known localization patterns, F-actin and PCNA. This choice was also motivated by the fact that interfering with actin or PCNA function would probably lead to visible phenotypes, conveniently showing side effects of chromobody expression. After injecting chromobody mRNA in zebrafish zygotes we could observe localized fluorescence at the expected sites where the respective antigens normally accumulate (Video 1). However, because of the expression variability and aspecific death in many embryos, we decided to switch to genetically-encoded expression systems. Therefore, by combining chromobodies and heat-shock or Gal4-dependent expression, we collected live imaging data supporting the actual binding of chromobodies to endogenous intracellular antigens in zebrafish, as confirmed by costaining with antibodies. With to the newly generated heat-shock and UAS transgenic lines we encountered some photobleaching problems, however we could detect actin and PCNA dynamics with high fidelity in different tissues or in the entire embryo, throughout embryonic development.
A very important question regards the impact of chromobodies in the context of a living embryo. Will they interfere with endogenous protein function? Strikingly, we observed that transgenic zebrafish embryos and larvae were morphologically normal throughout development and adult stages. They did not show aberrations in cell cycle progression (when monitoring the PCNA chromobody), nor they could alter the migration of lateral line primordium cells (in the case of the actin chromobody), even though it is known that interfering with actin dynamics at the leading edge of the primordium will perturb its migration (Xu et al., 2014). We sought a possible explanation for these observations. We detected fast recovery after photobleaching in HeLa cells stably expressing chromobodies. Importantly, the original localization of the antigen was immediately recapitulated by unbleached chromobodies, indicating that these molecules have elevated koff and kon, which imply a very transient antigen binding mode. We propose that the short binding half-life displayed by chromobodies prevents the emergence of global functional defects in living embryos.
In our work we show that chromobodies can be used as research reagents in living vertebrates, and substantially behave as already described in cultured mammalian cells. We introduce chromobodies as a complementary technique to more standard immunocytochemical and fluorescent-tag-based approaches. Chromobodies are advantageous because they allow live imaging without the need to directly modify or overexpress the protein of interest, approaches that can potentially produce artifacts. From the application side, chromobodies can be used as readout for phenotyping purposes or for high-throughput drug screening. As an example, the PCNA chromobody reports cell cycle progression just by displaying the localization dynamics of PCNA, which allows to differentiate single cell cycle stages and confers better resolution during S phase and S/G2 transition compared to FUCCI (Sakaue-Sawano et al., 2008).
Recent publications reported improved methods to clone and isolate nanobodies (Fridy et al., 2014), hence we hope that the high-throughput identification of novel binders will boost the availability of these reagents for basic research community. Although our results are preliminary because they focus only on two commercial chromobodies, we anticipate a widespread adoption of these promising reagents.
Featured article
Panza, P., Maier, J., Schmees, C., Rothbauer, U., & Sollner, C. (2015). Live imaging of endogenous protein dynamics in zebrafish using chromobodies Development, 142 (10), 1879-1884 DOI: 10.1242/dev.118943
References
De Meyer, T., Muyldermans, S. and Depicker, A. (2014). Nanobody-based products as research and diagnostic tools. Trends Biotechnol.32, 263–270. DOI: 10.1016/j.tibtech.2014.03.001
Fridy, P. C., Li, Y., Keegan, S., Thompson, M. K., Nudelman, I., Scheid, J. F., Oeffinger, M., Nussenzweig, M. C., Fenyö, D., Chait, B. T., et al. (2014). A robust pipeline for rapid production of versatile nanobody repertoires. Nat. Methods11, 1253–1260. DOI: 10.1038/nmeth.3170
Hamers-Casterman, C., Atarhouch, T., Muyldermans, S., Robinson, G., Hammers, C., Songa, E. B., Bendahman, N. and Hammers, R. (1993). Naturally occurring antibodies devoid of light chains. Nature363, 446–448. DOI: 10.1038/363446a0
Holliger, P. and Hudson, P. J. (2005). Engineered antibody fragments and the rise of single domains. Nat. Biotechnol.23, 1126–1136. DOI: 10.1038/nbt1142
Rothbauer, U., Zolghadr, K., Tillib, S., Nowak, D., Schermelleh, L., Gahl, A., Backmann, N., Conrath, K., Muyldermans, S., Cardoso, M. C., et al. (2006). Targeting and tracing antigens in live cells with fluorescent nanobodies. Nat. Methods3, 887–889. DOI: 10.1038/nmeth953
Sakaue-Sawano, A., Kurokawa, H., Morimura, T., Hanyu, A., Hama, H., Osawa, H., Kashiwagi, S., Fukami, K., Miyata, T., Miyoshi, H., et al. (2008). Visualizing Spatiotemporal Dynamics of Multicellular Cell-Cycle Progression. Cell132, 487–498. DOI: 10.1016/j.cell.2007.12.033
Schnell, U., Dijk, F., Sjollema, K. A. and Giepmans, B. N. G. (2012). Immunolabeling artifacts and the need for live-cell imaging. Nat. Methods9, 152–158. DOI: 10.1038/nmeth.1855
Swulius, M. T. and Jensen, G. J. (2012). The Helical MreB Cytoskeleton in Escherichia coli MC1000/pLE7 Is an Artifact of the N-Terminal Yellow Fluorescent Protein Tag. J. Bacteriol.194, 6382–6386. DOI: 10.1128/JB.00505-12
Xu, H., Ye, D., Behra, M., Burgess, S., Chen, S. and Lin, F. (2014). Gβ1 controls collective cell migration by regulating the protrusive activity of leader cells in the posterior lateral line primordium. Dev. Biol.385, 316–327. DOI: 10.1016/j.ydbio.2013.10.027
Hello! Read on for some fascinating stories of stem cell treatments – proven and unproven – and find out about a new website with great information to help you tell the difference.
The Hubrecht Institute have discovered a new way of introducing molecular tools into cells – read all about it in our latest research spotlight.
Plus a new French translation of the All About Stem Cells teaching tool, a freshly updated fact sheet on stem cells and spinal cord injuries, and some interesting events coming up.
Stem cell tourism: selling hope through unproven stem cell treatments – lessons from the X-Cell Center controversy
In recent years there has been a growing interest in so-called stem cell ‘tourism’ – where a person (often companied by their carer/family) travels to another country for a purported stem cell treatment that is not available in their home country. Many advertised treatments are clinically unproven, with little or no evidence for their safety and efficacy in specific conditions.
Interview with Graziella Pellegrini – Using stem cells to cure blindness
Professor Graziella Pellegrini is one of the principal scientists on the ground breaking, corneal repair system Holoclar®. Working throughout Italy over the past 27 years she is now based at the Centre for Regenerative Medicine “Stefano Ferrari” at the University of Modena and Reggio Emilia.We spoke to Graziella about developing Holoclar, what it means for regenerative medicine in Europe, and what’s next.
Interview with Michele De Luca – Epithelial cells and regrowing corneal epithelium
We spoke to Michele De Luca,director of the Centre for Regenerative Medicine “Stefano Ferrari” at the University of Modena and Reggio Emilia, about his work, the importance of collaboration, and his aspirations. He also shares some great advice for scientists starting out.
The International Society for Stem Cell Research (ISSCR) has launched a new website to help patients and their families make informed decisions about stem cell treatments, clinics and their health.
Stem Cells Australia’s Megan Munsie, chairperson of the ISSCR task force responsible for the website expansion, describes the revamped site as “a direct channel from researchers to the public.”
Researchers at the Hubrecht Institute and Utrecht University have developed a revolutionary and effective way of introducing molecular tools into cells. According to Dr. Niels Geijsen, who headed the research team, this discovery brings us one step closer to treating genetic diseases:
“The difficulty of treating genetic (inherited) diseases is that we, thus far, are unable to safely transport large therapeutic compounds, for example, proteins, into cells,” explains Geijsen. “With our new technology, we’ve found that we can do this very efficiently.”
New French translation of teaching tool: Tout sur les cellules souches
Un outil d’enseignement flexible qui introduit la recherche sur les cellules souches au moyen d’activités créatives en groupes et de discussions. Inclut des cartes d’activités imagées, des fiches de travail simples, des modèles de posters et un guide d’activités avec de nombreuses suggestions sur la façon d’utiliser le matériel.
28-30 May: Molecular Mechanisms of Neurodegeneration
This meeting will address the most important issues involved in protein toxicity in neurons, aiming at stimulating new and productive interactions among basic and clinical research groups. Oral presentations, plenaries and talks will provide participants with updated information that could be useful to researchers in the field of neurodegenerative disorders. Read more
This EMBO Conference will focus on the clinical translation of stem cell based research, current trials, pre-clinical work as the foundation for future ones and the ethical and economical impact of these novel therapies. Read more
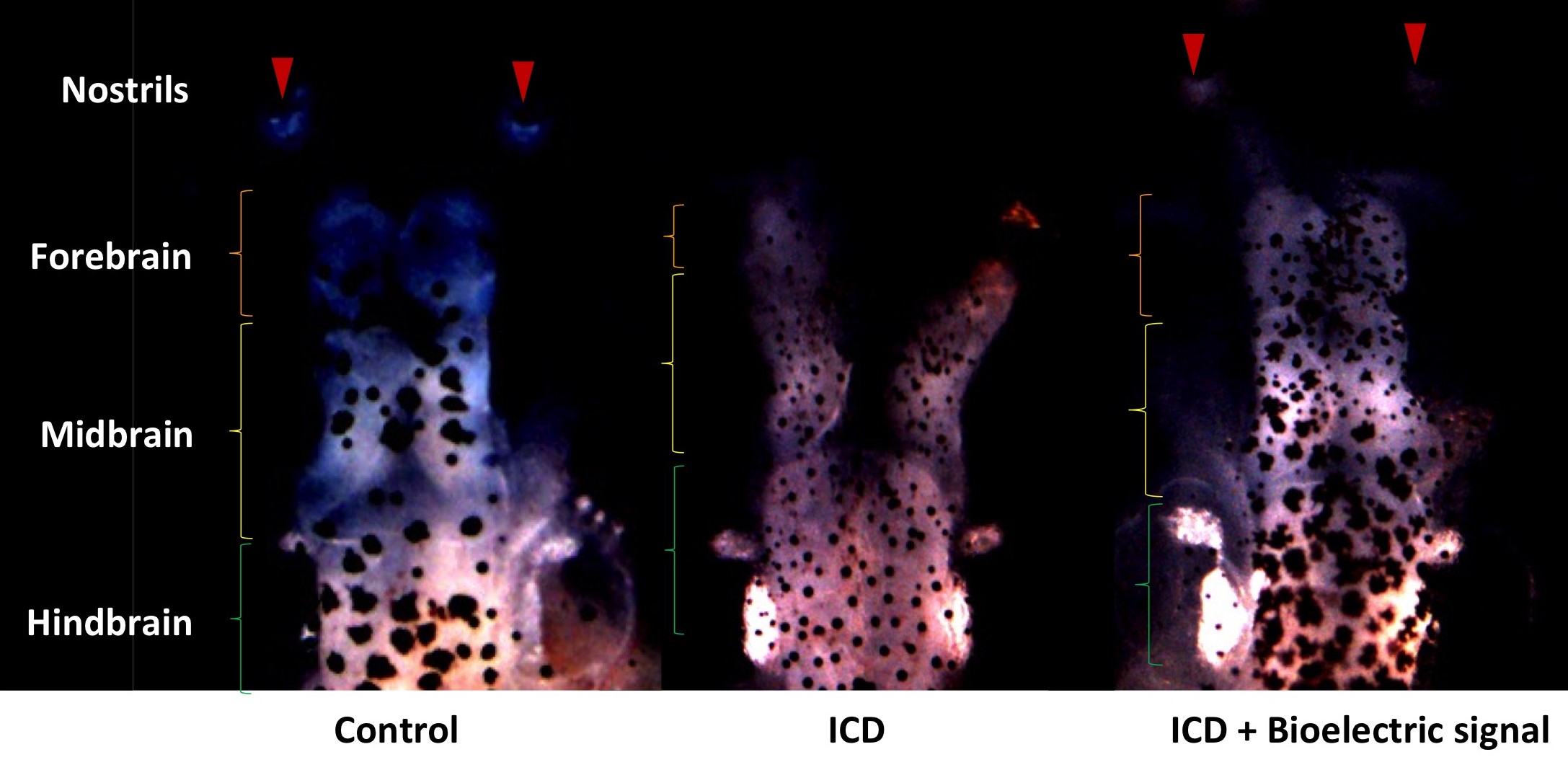
Work from Vaibhav Pai showing how manipulation of bioelectrical effects can rescue brain deficiencies caused by the introduction of Notch ICD
Trying to meet other young researchers in the Boston area interested in developmental biology can be surprisingly difficult. Despite the high density of scientists in a relatively small area, our day-to-day life exists mostly within our own labs; it’s arguable whether we interact enough with others in our own departments, never mind across departments and institutions.
After hearing about the Young Embryologist Network in the UK, it seemed that the perfect solution was to organize similar meetings this side of the pond. After lots of help from the YEN group in the UK, and kind support from Prof Marc Kirschner and the Department of Systems Biology at Harvard Medical School, on May 14th we had our first meeting, as advertised on the Node recently.
The first talk was due to be given by Dr. Stan Artap, a postdoc at Beth Israel Deaconess Medical Center in Peter Oettgen’s lab, talking about the “mouse heart” part of the meeting. However, in a great gesture that I hope will set the tone for our group going forward, Stan brought along his research assistant, Bryan Marsh, who is about to apply to graduate school in the area of developmental biology, to give the talk instead. Bryan, or our “embryological embryologist”/Very Young Embryologist, gave an excellent talk about the work he and Stan have carried out looking at the roles of the transcription factor ERG in cardiogenesis: early, in regulating BMP signaling during the specification of cardiogenic mesoderm into myocardial progenitors; and later in development, promoting endocardial-to-mesenchymal transition during cardiac cushion formation.
Left, Bryan Marsh presenting; right, an image of the eye by Vaibhav Pai.
The next talk was from Dr. Vaibhav Pai, a postdoc in Mike Levin’s Lab at Tufts University, talking about the role of membrane potentials in tissue specification in embryogenesis. Pai discussed his work in Xenopus manipulating ion flux to cause the formation of ectopic eyes in a Pax6-independent manner in all parts of the tadpole (Pai et al., 2012). He went on to show his recent work in rescuing Notch ICD-induced brain deformations by manipulating ion flux (Pai et al., 2015). His talk led to a very lively discussion, as a talk that demonstrates the role of biophysical forces in completely over-riding conventional gene regulatory network theories is wont to do!
Left, another of Vaibhav Pai’s eye images; right, Vaibhav Pai presenting.
The meeting format was very informal; people were free to grab pizza and soda as they needed, and the “20 minute talk, 10 minute question” format worked very well for the meeting (so well, that some participants suggested they would have enjoyed hearing a third speaker). Surveys were distributed to figure out what worked and what didn’t, and the feedback was very positive; people enjoyed the ability to hear talks from various speakers, and the opportunity to meet with other researchers from other institutions. In particular, everyone seemed very keen to return for the June meeting, which will feature Siyeon Rhee, a graduate student in Kimberley Tremblay’s lab at UMass Amherst, and Sam Morris, a postdoc in George Daley’s lab at Boston Children’s Hospital.
There are still some key challenges in getting the meeting up and running; representation was very Harvard Med School-heavy, and some of the questions going forward are where to hold meetings (in one location, or rotate around a few institutions?) and how to advertise the meeting effectively (if you’re reading this, and are interested, and now others who might be, please check out the website and get in touch/spread the word!) to build up a sustainable regular meeting. However, it was great to see such enthusiasm beyond just having free pizza, and to get together and talk science with other young embryologists.
A joint Biochemical Society / FEBS Focused Meeting, held to mark the retirement of Professor Robin Irvine, FRS. It will be organized by Len Stephens, Phill Hawkins, Colin Taylor and Peter Cullen. The meeting will begin on the afternoon of Tuesday 1 September, and conclude before lunch on Friday 4 September 2015. The link with Robin Irvine’s retirement has allowed us to attract the most eminent researchers in the field. There will be poster sessions from which early career speakers will be selected. The Colworth Medal lecture will be presented by Dr Helen Walden (Dundee). Sessions will be chaired by distinguished colleagues of Robin, including Professor Sir Michael Berridge, Professors Jim Putney and Bob Michell. There will be no evening sessions, but there will be an active programme of social events.
The ‘Biennial European Fly Neurobiology Conference’ is now more commonly known by its other name, ‘Neurofly’. And as of today, it has been about 7 months since the last Neurofly, which took place in October 2014 along a shore of the island Crete. Off the top of my head, the meeting was held at the Creta Maris Convention Centre. I do not recollect exactly how many Neurofly meetings have taken place (14, 15, 16?), and I am now guessing Neurofly 2014 took place sometime between the 10th and 19th of October. I do remember there was a good approval by show-of-hands on the last day of Neurofly to have the next meeting in Greece again.
Attending Neurofly was such as pleasure, I think, but now I find myself asking why it had been such a pleasure. I mentally draw out the landscape of a welcoming sandy beach… with a relaxing 5-star resort nearby… occupied by fellow friends and colleagues… who gathered in groups for meals and poster sessions, very much engaged in catch-up and scientific discussions… We listened to lectures and short talks in an air-conditioned conference hall. It is always fun to communicate our own research and to listen to those of others, yet calling forth detailed contents of each talk purely on memory is naturally impossible. I know the opening afternoon of Neurofly was graced with 2 plenary lectures, of which the first speaker discussed the complexity of Notch signalling. The second speaker discussed the [daresay] necessary biology of forgetting, in which dopamine signalling is involved.
More accurately, there has been 15 Neurofly to date and the 15th took place on the shores of Hersonissos on Crete from Sunday, 5th October, to Thursday, 9th October 2014. A welcoming first day greeted attendees at the Creta Maris Convention Centre with two plenary lectures in the late afternoon. Spyros Artavanis-Tsakonas had expertly navigated the convoluted complexity of the Notch signaling pathway while Ron Davis, lest we forget, took us back in time to 1974, just over 40 years ago, to explore the milestones in Drosophila memory research since. The history was followed on by Davis putting forward that the biology of forgetting is necessary, being also a form of ‘homeostasis’ after a learning event, and that dopamine signaling is key in memory acquisition and forgetting.
Amongst the talks in the Development and Differentiation session, Angela Giangrande seeks to decode the molecular interactions between transcriptional fate determinants which drive stable glial cell fate. Glia depends on the glial cell missing (Gcm) transcription factor and the reverse polarity (Repo) homeodomain protein to dictate neural stem cells for choice of either neuronal or glial identity. The Giangrande research team demonstrated an existence of a temporal sequence of regulatory loops. In the earlier part of the sequence, Gcm controls its own level of expression until the threshold necessary to trigger gliogenesis is reached. As soon as this threshold is achieved, a Gcm target starts to degrade Gcm itself to allow a stable differentiation of glia. Repo acts together with Gcm such that the level of the former also ascends and reaches threshold after Gcm’s peak. Then Repo acts by inhibiting Gcm to influence the outcome of cell differentiation into glia.
Another interesting talk was given by Jean-Maurice Dura, who expanded the Wnt5/Derailed-mediated axon guidance mechanism from more familiar grounds and into fascinating new landscape with its descriptions of ligand capturing and receptor shedding during brain development. A personal highlight was the work of Adel Dunayevskyy and Adi Salzberg from the Technion-Israel Institute of Techology in Israel. Salzberg talked about examining an enigmatic phenomenon of numbers in developmental biology. A cluster of five proprioceptive chordotonal organs contains five neuron, five scolopale cells, five cap cells, five ligament cells…, but only two cap attachment (CA) cells. Indeed, it must be a case of ‘five minus three’ CA cells for proper morphogenesis. For even when developmental apoptosis is perturbed, the developing organ insists to eliminate unwanted excess cells and it does so by switching on autophagy.
On day 3 of Neurofly, there was a window of opportunity between talks and dinner time for attendees to visit Knossos Palace, a relic from the Bronze Age and an archaeological site amidst beautiful hilly surroundings. The day eventually led to scientists and accompanying guests feeding on the gastronomic delights of Cretan dishes at the Cretan Thematic Park.
During dinner, I took interest in the history of Neurofly, specifically on how it started. This sparked a discussion on past Neurofly events at the dinner table. The scientists sitting with me on this conversation were 2 of the 4 host organisers, Efthimios Skoulakis and Maria Monastirioti, and Carsten Duch. We didn’t get to find out how and why Neurofly started, but we did gather a list of past Neurofly locations. Here is the initial list which we came up with:
?
?
1990 – ?
1992 – Glasgow, UK
1994 – Regensbourg, Germany
1996 – Würzburg, Germany
1998 – Warwick, UK
2000 – ?
2002 – Alicante, Spain
2004 – Dijon, France
2006 – Würzburg, Germany
2008 – Manchester, UK
2010 – Belgium
2012 – Padua, Italy
2014 – Crete (Hersonissos), Greece
After the meeting, I found out that everyone was fairly good in recollecting past Neurofly locations. I refined the list through an update of the year-location pairing, and I found out that the 1st to 11th meeting were known in their time as the ‘European Meeting on the Neurogenetics of Drosophila’. Here is the updated list:
1986 – Simonswald, Germany
1988 – ?
1990 – Saint-Rémy, France
1992 – Glasgow, UK
1994 – Montpellier, France
1996 – Regensbourg, Germany
1998 – Warwick, UK
2000 – Alicante, Spain
2002 – Dijon, France
2004 – Neuchatel, Switzerland
2006 – Leuven, Belgium
2008 – Würzburg, Germany
2010 – Manchester, UK
2012 – Padua, Italy
2014 – Crete (Hersonissos), Greece
The venue of the second Neurofly remains unknown. Should anyone uncover the mysterious location, please respond by commenting below.
It has been a pleasure for me to report on Neurofly 2014. If you’re interested in attending the next Neurofly meeting, keep an eye on the upcoming meetings webpage hosted by FlyBase.
Stem cells play crucial roles in development as well as tissue homeostasis, repair, and regeneration, and their dysregulation is involved in diseases and aging of the tissues. The stem cell is defined as a cell that has the ability to self-renew and also to produce differentiated progeny for a long-term. Yet, stem cells require other functional properties to regulate a variety of biological processes.
Epidermal keratinocyte stem cells are one of well-characterized somatic stem cells in human and successfully applied for autologous transplantation of epidermal sheets onto patients with extensive burns and genetic disorders. However, keratinocyte stem cells are eventually converted into keratinocytes with more restricted proliferative capacity. So we lose the stem cells during serial cultivation, and as a result, sometimes fail the transplantation. Therefore, a noninvasive quality control of the stem cell culture is essential for successful regenerative medicine.
We have previously found that cultured human keratinocyte stem cells and transient amplifying (TA) cells differ in their actin filament organization. This causes the two distinct modes of epidermal growth factor (EGF)-induced colony dynamics in colonies formed by keratinocyte stem cells and TA cells1. So, we decided to explore the relationship between spontaneous cell motion and proliferative capacity in cultured human keratinocytes to identify the stem cells noninvasively and in situ.
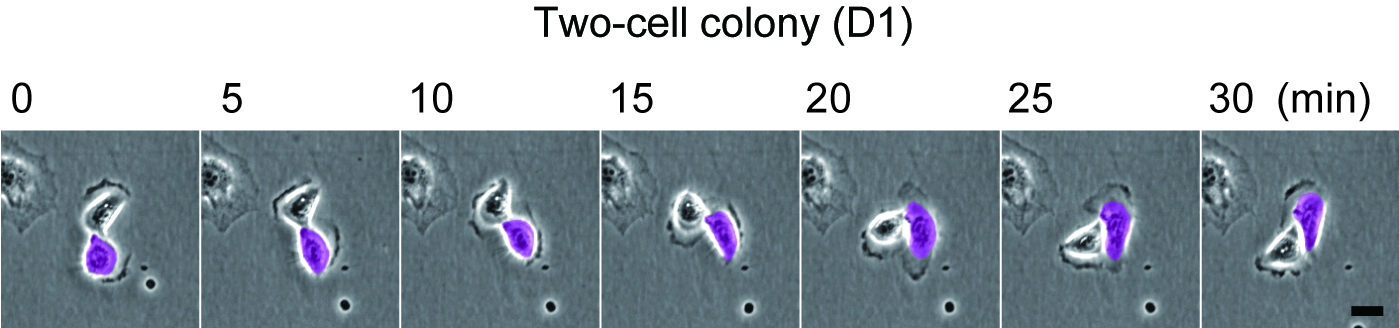
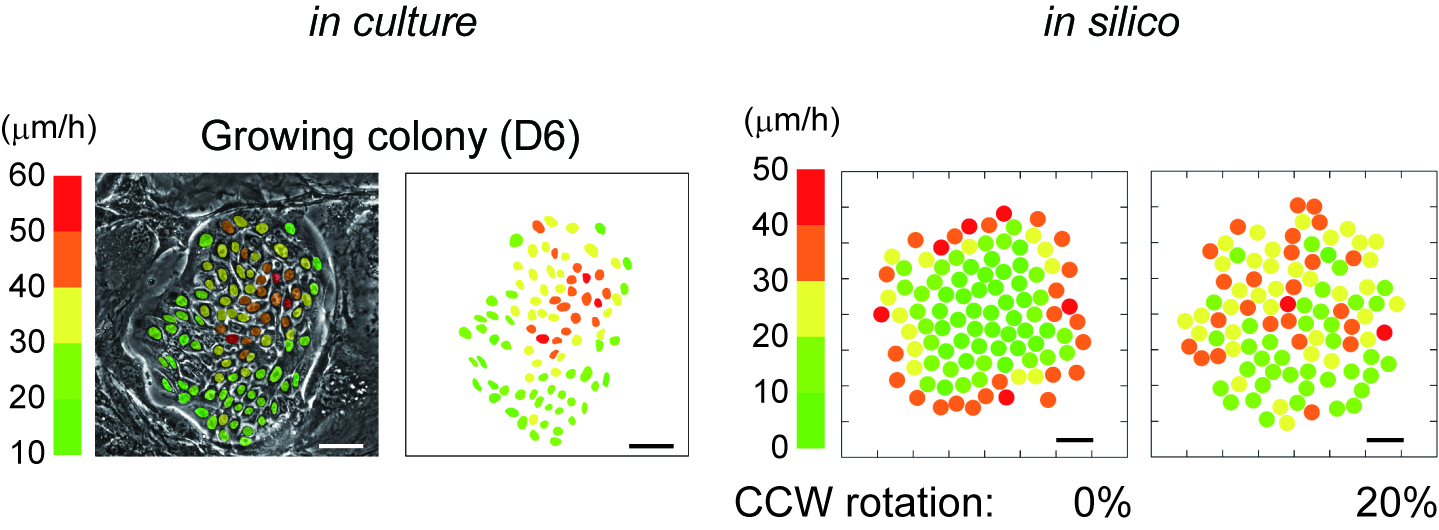
As described in our resent paper2, we found that keratinocytes with significant proliferative capacity displayed a unique rotational motion at the two-cell stage colony which is formed by a single cell division of individual keratinocytes (Figure 1). We then simulated the interaction of a hundred rotating keratinocytes with physics of multiparticle systems and proved that this rotational motion of cells gives rise to collective motion of keratinocytes in the colony (Figure 2). This in silico experiments also revealed the possibility of predicting a keratinocyte stem cell colony by only measuring cell locomotion speed. We then validated this prediction with a combination of time-lapse observation and clonal analysis with human keratinocyte stem cell culture system, and finally, we could identify human keratinocyte stem cell colonies with a noninvasive procedure.
Rotational motion of a two-cell colony of human epidermal keratinocytes.
Figure 1 Rotational motion of a two-cell colony oh human keratinocytes.
Human epidermal keratinocytes were seeded on one fourth of normal number of feeder cells. The two-cell colonies of keratinocytes (paired keratinocytes) were observed 1 day later. Bar, 20 um.
In silico reconstruction of keratinocyte stem cell colony dynamics.
Figure 2 In silico reconstruction of keratinocyte stem cell colony dynamics.
Left panel shows that distribution of locomotion speed of keratinocytes in a growing colony after 6 days cultivation. Each color indicates different locomotion speeds that were calculated with 3 hours time-lapse observation. Bar 100 um. Right panel shows that distribution of locomotion speed of keratinocytes in a growing colony in silico. The speed of each cells was calculated by a simulation experiment based on our modeling. Bar, 50 um.
We also found that alpha6 integrin, which is a known marker of keratinocyte stem cells, is required for rotational and collective motion of keratinocytes. alpha6 integrin is also highly expressed at the leading edge of migrating epidermis during wound repair. Although the mechanism of the rotational motion associated with growth capacity remains unclear, motile epithelial cells could retain the significant proliferative capacity, or a population of migrating cells might contain stem cells. It seems to be involved in development as well as tissue repair and regeneration.
2. Nanba, D., Toki, F., Tate, S., Imai, M., Matsushita, N., Shiraishi, K., Sayama, K., Toki, H., Higashiyama, S., & Barrandon, Y. (2015). Cell motion predicts human epidermal stemness The Journal of Cell Biology, 209 (2), 305-315 DOI: 10.1083/jcb.201409024
This is a news item which was first posted on thebsdb.org site. Please, note that not all items will be duplicated on The Node. To ensure you stay informed, please, take a minute to subscribe for email notifications on the bsdb.org site: simply enter your email address in the 3rd item of the side bar. Be ensured that the amount of emails sent to you will usually not exceed one per week or fortnight.
—————————–—————————–
The lectures of the two BSDB medal award winners announced on the 2015 Spring Meeting are now available on YouTube:
the Waddington Medal lecture by Lewis Wolpert (UCL, London) who presented a talk about his seminal discoveries of concepts of positional information, and
the Beddington Medal lecture by John Robert Davis who presented a talk entitled “Intercellular forces orchestrate cell repulsion and embryonic pattern formation“.
These two talks are embedded below. Information about all other awardees, links to interviews performed by The Node, and a meeting report can be found in an earlier post which has been updated accordingly.

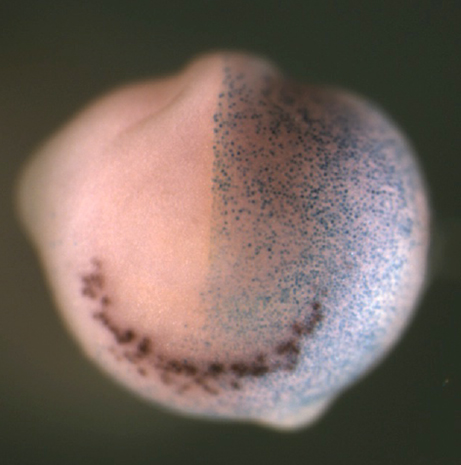 Neuroblastoma (NB) is a paediatric form of cancer derived from the sympathetic nervous system. Recent genome-wide sequencing data suggest that often NB does not have a clear genetic cause, leading the authors to hypothesize that NB results from aberrations of normal development. To test this hypothesis, Anna Philpott’s group used a population of anteroventral noradrenergic (AVNA) cells from Xenopus embryos. These cells share several features with mammalian sympathetic neurons, including the expression of noradrenergic-associated genetic markers such as the achaete-scute complex-like 1 (Ascl1) gene, which encodes a transcriptional driver of neurogenesis. By comparing AVNA and NB cells, the authors found that, whereas Ascl1 is only transiently expressed in AVNA cells, it is aberrantly maintained in NB, where it is phosphorylated on multiple serine-proline sites. The authors then show that differentiation of AVNA cells is enhanced by dephosphorylated Ascl1. Moreover, this process is inhibited by experimental manipulations of NB-associated genes, but, interestingly, dephosphorylation of Ascl1 is able to overcome this inhibition. This work demonstrates that Xenopus AVNA cells represent a unique system to study sympathetic nervous system development and its relationship to NB. Moreover, it suggests that Asc11 phosphorylation might promote stalled differentiation leading to NB, thus identifying a potential target for therapeutic purposes. Read the paper here (Open Access) and the authors’ Node post here.
Neuroblastoma (NB) is a paediatric form of cancer derived from the sympathetic nervous system. Recent genome-wide sequencing data suggest that often NB does not have a clear genetic cause, leading the authors to hypothesize that NB results from aberrations of normal development. To test this hypothesis, Anna Philpott’s group used a population of anteroventral noradrenergic (AVNA) cells from Xenopus embryos. These cells share several features with mammalian sympathetic neurons, including the expression of noradrenergic-associated genetic markers such as the achaete-scute complex-like 1 (Ascl1) gene, which encodes a transcriptional driver of neurogenesis. By comparing AVNA and NB cells, the authors found that, whereas Ascl1 is only transiently expressed in AVNA cells, it is aberrantly maintained in NB, where it is phosphorylated on multiple serine-proline sites. The authors then show that differentiation of AVNA cells is enhanced by dephosphorylated Ascl1. Moreover, this process is inhibited by experimental manipulations of NB-associated genes, but, interestingly, dephosphorylation of Ascl1 is able to overcome this inhibition. This work demonstrates that Xenopus AVNA cells represent a unique system to study sympathetic nervous system development and its relationship to NB. Moreover, it suggests that Asc11 phosphorylation might promote stalled differentiation leading to NB, thus identifying a potential target for therapeutic purposes. Read the paper here (Open Access) and the authors’ Node post here.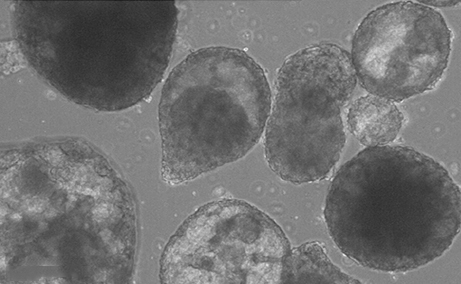 Duchenne muscular dystrophy (DMD) is a genetic muscular disorder characterised by progressive muscular weakness and wasting, with cardiac complications (such as dilated cardiomyopathy) and respiratory muscle failure arising in the late stage. To investigate mechanisms of dilated cardiomyopathy in DMD, Lei Yang and collaborators derived cardiomyocytes (CMs) from DMD-patient-specific induced pluripotent stem cells (iPSCs). Compared to control cells, DMD iPSC-CMs exhibited elevated cytosolic calcium, mitochondrial damage and increased cell apoptosis. To further dissect the mechanisms underlying these alterations, the authors performed transcriptional and translational analyses and identified a mitochondrially initiated molecular cascade – which involves CASPASE3 (CASP3) activation – as being responsible for the increased apoptosis in DMD iPSC-CMs. Notably, the application of the membrane sealant Poloxamer 188 could prevent calcium overload and CASP3 activation, significantly reducing apoptosis in these cells. Thus, the authors established a useful in vitro system to disclose mechanisms of cardiomyopathy in DMD and to identify molecular targets that could be pharmacologically manipulated. Read the paper here (Open Access).
Duchenne muscular dystrophy (DMD) is a genetic muscular disorder characterised by progressive muscular weakness and wasting, with cardiac complications (such as dilated cardiomyopathy) and respiratory muscle failure arising in the late stage. To investigate mechanisms of dilated cardiomyopathy in DMD, Lei Yang and collaborators derived cardiomyocytes (CMs) from DMD-patient-specific induced pluripotent stem cells (iPSCs). Compared to control cells, DMD iPSC-CMs exhibited elevated cytosolic calcium, mitochondrial damage and increased cell apoptosis. To further dissect the mechanisms underlying these alterations, the authors performed transcriptional and translational analyses and identified a mitochondrially initiated molecular cascade – which involves CASPASE3 (CASP3) activation – as being responsible for the increased apoptosis in DMD iPSC-CMs. Notably, the application of the membrane sealant Poloxamer 188 could prevent calcium overload and CASP3 activation, significantly reducing apoptosis in these cells. Thus, the authors established a useful in vitro system to disclose mechanisms of cardiomyopathy in DMD and to identify molecular targets that could be pharmacologically manipulated. Read the paper here (Open Access).
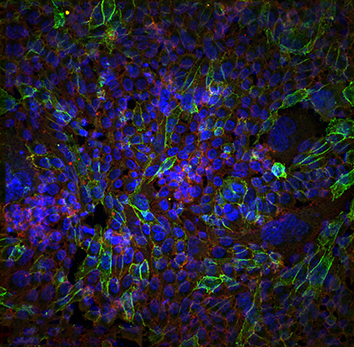 Epithelial-to-mesenchymal transition (EMT) is an integral developmental and physiological process, but can also be utilised by cancer cells at the initiation of metastasis. A requirement for EMT is the post-translational removal of adhesion proteins from the plasma membrane. Here (p. [164426]), Lisa Taneyhill and Rangarajan Padmanabhan study cadherin-6B (Cad6B) internalisation to elucidate the mechanisms of EMT in chick cranial neural crest cells. The authors found that in neural crest cells that are initiating EMT, Cad6B was detected in cytosolic puncta that were endocytic, rather than exocytic, in nature. They then identified two intracellular motifs that were potentially important for regulating Cad6B internalisation. Mutating the p120-catenin-binding (EED) motif, but not the dileucine (LI) motif, significantly increased Cad6B internalisation, supporting the idea that Cad6B is removed from the plasma membrane through endocytosis. However, although Cad6B colocalised with clathrin, the colocalisation was not exhaustive, suggesting that an additional mechanism is involved in Cad6B internalisation. Therefore, the authors used an array of pharmacological treatments to show that Cad6B was removed from the plasma membrane through both endocytosis and macropinocytosis, and that both of these processes depended on dynamin. This study demonstrates that EMT and neural crest migration require Cad6B internalisation through endocytosis and macropinocytosis. Read the paper here.
Epithelial-to-mesenchymal transition (EMT) is an integral developmental and physiological process, but can also be utilised by cancer cells at the initiation of metastasis. A requirement for EMT is the post-translational removal of adhesion proteins from the plasma membrane. Here (p. [164426]), Lisa Taneyhill and Rangarajan Padmanabhan study cadherin-6B (Cad6B) internalisation to elucidate the mechanisms of EMT in chick cranial neural crest cells. The authors found that in neural crest cells that are initiating EMT, Cad6B was detected in cytosolic puncta that were endocytic, rather than exocytic, in nature. They then identified two intracellular motifs that were potentially important for regulating Cad6B internalisation. Mutating the p120-catenin-binding (EED) motif, but not the dileucine (LI) motif, significantly increased Cad6B internalisation, supporting the idea that Cad6B is removed from the plasma membrane through endocytosis. However, although Cad6B colocalised with clathrin, the colocalisation was not exhaustive, suggesting that an additional mechanism is involved in Cad6B internalisation. Therefore, the authors used an array of pharmacological treatments to show that Cad6B was removed from the plasma membrane through both endocytosis and macropinocytosis, and that both of these processes depended on dynamin. This study demonstrates that EMT and neural crest migration require Cad6B internalisation through endocytosis and macropinocytosis. Read the paper here.
 Spontaneously beating 3D ‘heart’ structures can be developedin vitro from larval zebrafish using a novel, fast and inexpensive method that could be employed in ecotoxicology and biomedical safety testing. Read the paper here.
Spontaneously beating 3D ‘heart’ structures can be developedin vitro from larval zebrafish using a novel, fast and inexpensive method that could be employed in ecotoxicology and biomedical safety testing. Read the paper here. (2 votes)
(2 votes)




 (No Ratings Yet)
(No Ratings Yet)