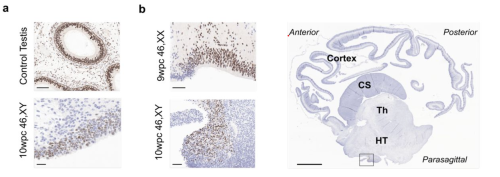
It’s challenging to sustain a pregnancy when food is short, or conditions are otherwise tough. That’s why many mammalian embryos can postpone their growth to get through periods of environmental stress and then re-enter development when conditions improve. This stalling of development is known as embryonic diapause, and understanding the mechanisms behind it might help improve infertility treatments, such as embryo freezing. Now, researchers at the Center for Excellence in Brain Science and Intelligence Technology, the Chinese Academy of Sciences in Shanghai, China, have discovered how nutrient depletion is sensed by embryos growing in hungry mouse mums to induce diapause. They publish their study in the journal Development on 11 April 2024.
A mouse embryo (blastocyst) that has paused its development due to nutrient depletion. Image credit: Jiajia Ye.
Lack of food is a known trigger of embryonic diapause, but it has not been clear how nutrient depletion in the mother’s diet is sensed by the embryo. “Seasonal starvation is one of the universal environmental stresses in nature,” explained Professor Qiang Sun, who led the study. “However, the regulatory process of diapause in early-stage embryos is not fully understood. So, we decided to examine whether nutrient deprivation induces embryonic diapause.”
By comparing hungry and well-fed pregnant mice, the team discovered that embryos in the hungry mice did not implant into the uterus and their growth paused at an early timepoint, when the embryo comprises a hollow ball of cells called the blastocyst. These embryos remained viable and could start developing again when transplanted into a well-fed mother.
To work out which nutrients were important to induce diapause, the researchers grew early-stage mouse embryos in dishes that contained different nutrients. They found that embryos grown in dishes lacking protein or carbohydrates paused their development, whereas the embryos exposed to normal nutrient levels did not stall and kept on developing. The scientists then went on to reveal that nutrient sensors in the embryo can detect drops in protein or carbohydrate levels, which triggers the entry into diapause.
The finding that embryos grown without protein or carbohydrates can pause their development means that they can survive longer in the lab. In the future, this finding might lead to improvements in fertility treatments, which currently include approaches such as embryo freezing. “We think our study can inspire the development of new methods for human embryo preservation,” said Professor Sun. “Embryo cryopreservation is a widely used approach, but there is still no consensus on when cryopreserved embryos can be thawed and transferred into the uterus. Many clinical studies have shown that traditional frozen embryo transfer can increase the risk of problems during pregnancy. Therefore, it is necessary to develop alternative methods to preserve embryos.”
Studies focusing on diapause may even have long-term implications for cancer treatments. “Dormant cancer cells which persist after chemotherapy resemble the diapaused embryos,” said Professor Sun. “Consequently, we hypothesize that delving into the mechanism of diapause may have positive implications for cancer treatment and decreasing the chances of relapse.”
Jiajia Ye, Yuting Xu, Qi Ren, Lu liu and Qiang Sun (2024). Nutrient deprivation induces mouse embryonic diapause mediated by Gator1 and Tsc2. Development, 151, dev202091 doi: 10.1242/dev.202091
This series of interviews features principal investigators (PIs) within the first five or so years of establishing their own research group. Through these conversations, Development aims to illustrate that there is not a ‘one-size-fits-all’ approach to securing an independent position and setting up a research programme. Discussing the challenges and difficulties new PIs have overcome and highlighting the best moments will hopefully offer encouragement to other ECRs and stimulate discussion around the career path of a developmental biologist.
Click on the pins to read the interviews:
Collage of all 30 interviewees in the ‘Transition in development’ series. (No Ratings Yet) Loading...
The lab is located in the southern part of the city of Lyon, France on the campus of the Ecole normale supérieure (ENS). It is one of the CNRS research groups at the Institute of Functional Genomics of Lyon (IGFL), which focuses on integrating developmental, physiological and evolutionary perspectives to study complex biological systems at the organism level.
Research summary
Our group studies the morphogenesis of brown algae. These organisms, some of which can grow to 45 m long, have evolved independently of plants and animals. We work on identifying the strategies that these organisms have adopted to establish their body plans at the embryogenesis stage. In particular, right now we are exploring which mechanisms control the position and the orientation of cell division planes. When, where and how do cells grow and divide is our current obsession!
This question is even more challenging when applied to our favourite models: brown algae are largely understudied multicellular organisms that possess unique cellular components – in terms of cell wall composition, vesicular and cytoskeleton organisation, pigments – resulting from their distinctive evolutionary pathway.
Lab roll call
Marie Zilliox: I’m a post-doc with a PhD in zebrafish development, and my mission is to monitor the growth of brown algal embryos in real time and in 3D using light-sheet and multi-photon microscopy techniques to determine whether cell shape controls the orientation of cell division. The challenge lies in mastering long-term time-lapse microscopy in 3D and in culture conditions compatible with the natural environment as well as with the presence of pigments that diffract light.
Tanguy Dufourt: I’m a first-year PhD student. I hold a Master’s degree in Systematic Evolution and Paleontology (MNHN Paris), and I am now studying which intracellular mechanisms are involved in the orientation of cell divisions in brown algal cells. I aim to model the displacement of centrosomes during the cell cycle by using software based on the forces generated by and applied to dynamic microtubules. First, I need to adapt this software to the cell biology of brown algae: cuboid cell shapes, presence of chloroplasts and vacuoles, throughout the cytoplasm, etc.
Alexis Martel: I have a Master’s in Marine Biology (Toulon University, France), more specifically in biotic interactions and anthropogenic disturbances in marine environments. I work here as a lab manager. In the past 6 months, I have set up most of the lab equipment and the cultures of the brown algal models of the team at IGFL. Now I am developing new microfluidics protocols for the forthcoming team projects involving this technique.
Bernard Billoud: I am a lecturer with a 30-year university career in bio-informatics, and a researcher working on modelling and simulation of algal development. My role is to develop mathematical and mechanical models that can account for the tissue patterning in brown algae. As a member of a team where most of the members are involved in experimental work on the bench, I can assess the time and technicality required to obtain the quantitative data on living organisms that I need to validate the models.
Bénédicte Charrier: I am the leader of the team. As a former land plant biologist, I have been studying the development of brown algae for the past 20 years. I come from a molecular and genetic background, and I am now studying the role of mechanical forces in the process of morphogenesis. Interdisciplinarity combining cell biology with mathematics and physics is what I need to answer my questions.
Favourite technique, and why?
Bénédicte: I love penetrating the microscopic world! Beyond observing live or fixed, labelled or even unlabelled cells, I am now captivated by microinjection. It provides me with an “intimate” moment spent with the algae. Microinjection requires lengthy preparation, but when the time has come for the needle to touch the cell surface, then the conversation starts. And in this specific context, different cells of brown algae reveal their lot of unexpected responses!
Apart from your own research, what are you most excited about in developmental and stem cell biology?
Bénédicte: I’d like to understand the complexity of biological systems. How do cells co-ordinate themselves or work together to form an organism? How do the different molecules that make up a cell interact to produce a functional living cell? What are their priorities and tolerances (plasticity, robustness)? What are their molecular and chemical bases? What role does self-organisation play? I like to think that the mechanisms of life are based on simple mechanistic rules, perhaps made up of complex interactions at the chemical/molecular level.
How do you approach managing your group and all the different tasks required in your job?
Bénédicte: I would like to say that I’ve taken part in training courses organised by my employer to help me manage all the tasks I have to do as well as possible, but that’s not the case! We’re a small team with a large proportion of young scientists in training. The belief that we’re all highly committed and motivated by our work is undoubtedly the cement to building trust among everyone on the team. My favourite time of the week is when I’m fed fresh results, straight from the oven!
What is the best thing about where you work?
Marie: The mutual support between the people from different research teams at the IGFL and also between the different laboratories and facilities located in Lyon (e.g. the Reproduction and Development of Plants laboratory, the PLATIM Imaging Core Facility) is a real help and relief especially when setting up a new laboratory, which is the case for the Charrier lab, which recently joined IGFL.
Tanguy: A good point of the lab is that the infrastructure is recent and the office is very close to the lab.
Alexis: Our laboratory is affiliated with the ENS, which is a renowned research centre with a vibrant and stimulating environment. The lab itself is recent and bright, so it’s quite pleasant to work in. The different teams work on very varied and interesting subjects.
Bernard: The lab is a very stimulating environment, where people work very seriously in a relaxed and enthusiastic atmosphere.
Bénédicte: The IGFL is an open-minded institute, with a wide variety of model organisms and scientific questions. You would think that this diversity would be unsettling or uncomfortable on a day-to-day basis when it comes to technical difficulties or model-specific issues, but it’s not. Open-mindedness is the DNA of the IGFL! What’s more, the institute is currently broadening its approach to developmental biology and recruiting new teams from different backgrounds. The fact that the IGFL is located on the ENS Lyon campus means that there are plenty of seminars by world-class scientists. The campus is quite cosy, but still offers a wide range of opportunities to develop my project in excellent conditions.
What’s there to do outside of the lab?
Marie: The laboratory is perfectly situated close to the Lyon city centre, where there are plenty of restaurants and bars to suit all tastes, from a cozy café to a night of dancing. Lyon is full of things to do, with an exhibition or concert every weekend. Lyon’s location is also ideal for getting to cities such as Annecy, Marseille, Paris and Geneva in less than 2 hours, which is perfect for weekend getaways.
Tanguy: In Lyon there are a lot of cultural places, like concert halls or museums.
Alexis: Lyon is a very pleasant city. The “Vieux-Lyon”, which is the old quarter in the city centre is a pleasant place to wander around. Also “le Parc de la Tête d’Or” is a peaceful green city park, where I like to go for walks.
Bernard: Lyon has all practical and cultural facilities of a big city, with relaxing natural sites nearby.
Bénédicte: The Alps being less than 2 hours’ drive away is a major advantage when you like to be outdoors, in the mountains. The Mediterranean Sea isn’t that far away either. Lyon is also actively developing its cycling infrastructure, with greenways available in the near future. This is a considerable advantage when, like me, stopping taking the car to work becomes a daily priority.
In this SciArt profile, we meet Maja Mielke, who is doing a PhD in functional morphology and enjoys making nature-inspired drawings.
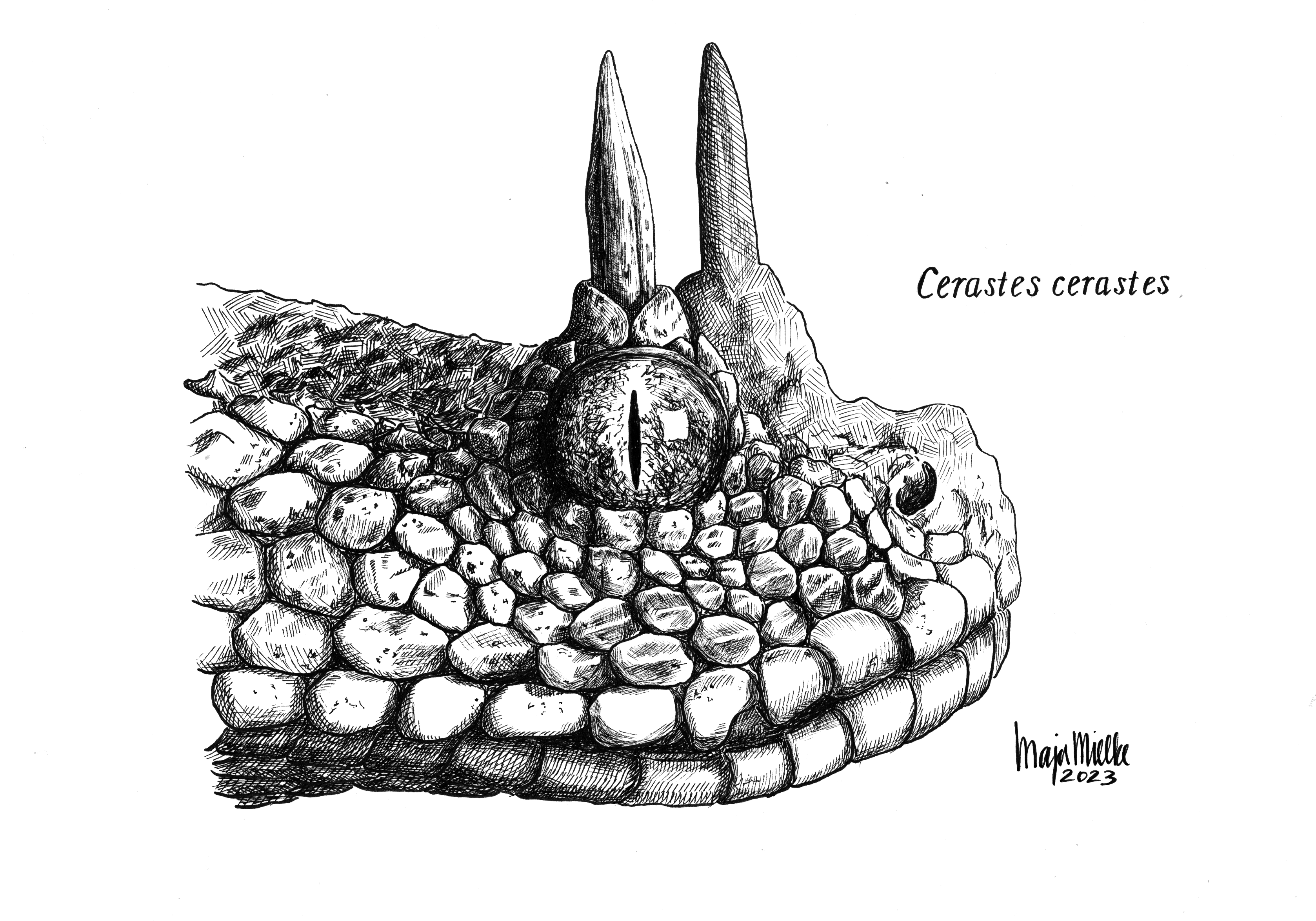
Saharan horned viper (pen and ink). I used a photo by National Geographic photographer Joel Sartore as reference for this illustration.
Can you tell us about your background and what you work on now?
Currently, I’m pursuing a PhD in functional morphology at the University of Antwerp (Belgium), but originally, I’m from Germany. I did my bachelor’s in Molecular Biophysics in Berlin but switched to Biology & Evolution for my master’s because I wanted to study animals rather than molecules. This way, I discovered my fascination for the functional morphology and biomechanics of vertebrates. I studied squirrels for my master’s thesis and switched to birds for my PhD. I’m exploring how they move their beak while processing, cracking, and husking seeds.
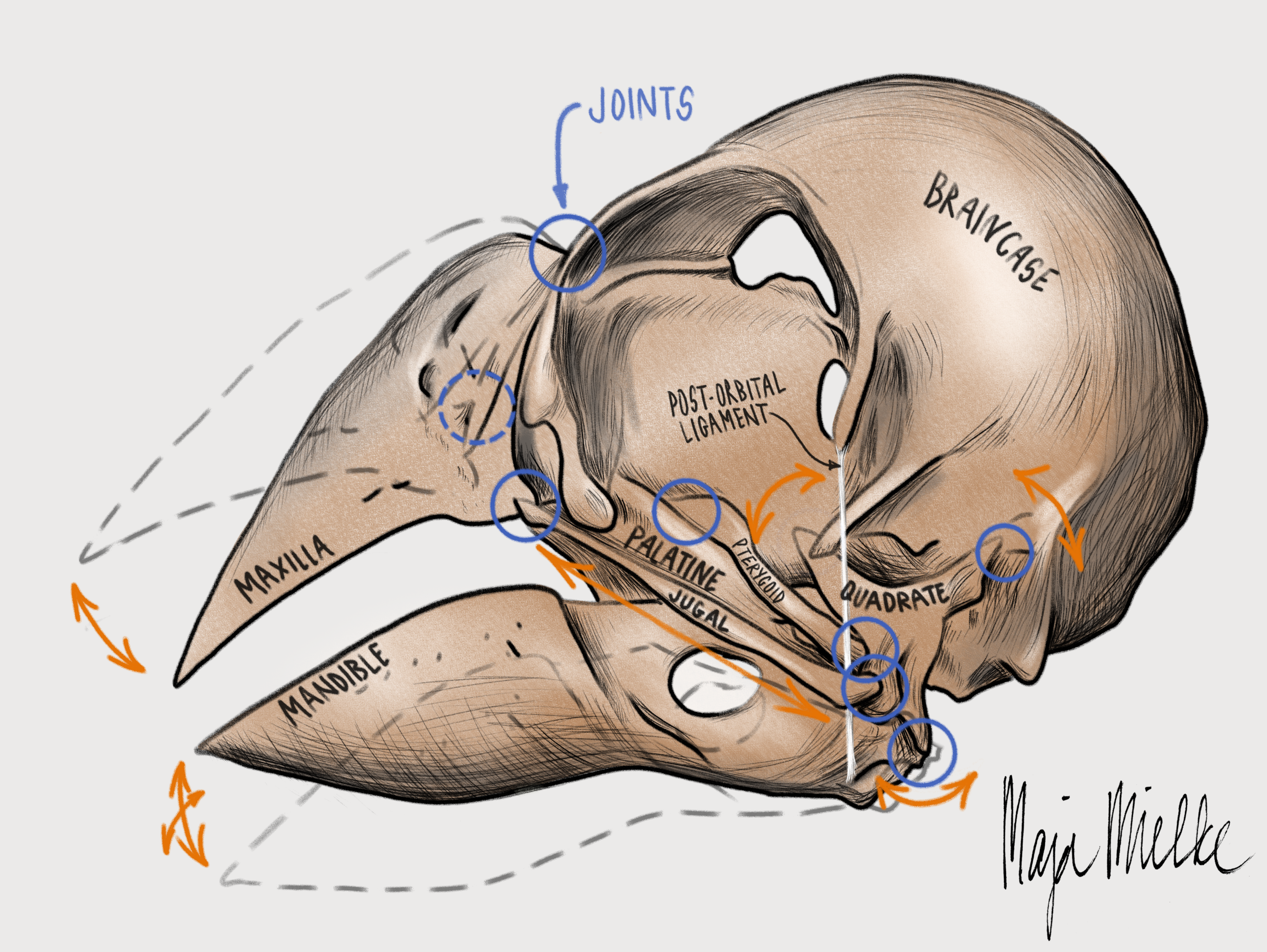
Skull of a Java finch (digital illustration made in Krita). I rarely do digital illustrations, but the flexibility that comes with working digitally comes in handy for work-related illustrations. I made this drawing to pitch my PhD project to a research foundation. It worked! They are now funding my PhD.
Were you always going to be a scientist?
Pretty much, yes. Especially the natural sciences have always sparked my interest and fascination for the world around us. Becoming a scientist was the logical step after finishing school. The hardest part was to choose a field of study! But the interdisciplinary Biophysics bachelor program was the perfect fit at that time.
And what about art – have you always enjoyed it?
I have practiced and enjoyed art a lot during my school years. But once I started my bachelor studies, I was so occupied with lectures, practicals, and exams that there was hardly any room for creating art. Only during my masters did I attempt to re-integrate an art practice into my life. Unfortunately, while managing work, family, and other hobbies, I still haven’t managed to practice art on a regular basis until now. But whenever I finish a piece, it makes me really happy.
Great horned owl (pen and ink). This owl was one of the first more elaborate pieces I did with pen and ink. I used a photo by National Geographic photographer Joel Sartore as a reference for this illustration.
What or who are your most important artistic influences?
First, my art is primarily inspired by nature and my fascination for the animal world. That determines my ideas on what to illustrate in the first place. Second, I’m inspired by the work of both professional artists (like Ben Rothery, Alphonso Dunn, Denise Soden, and Raoul Deleo) and fellow hobby artists that share their work online. Studying their art influences my own artistic approaches and inspires me a lot. Third, my art is highly influenced by the work of wildlife photographers, whose beautiful work I use as references for my drawing practice. Last, I’ve been highly influenced by drawing and painting courses that I attended during my life, be it a painting class for children during primary school, more advanced naturalistic drawing classes during high school, or a scientific illustration course I attended two years ago during my PhD.
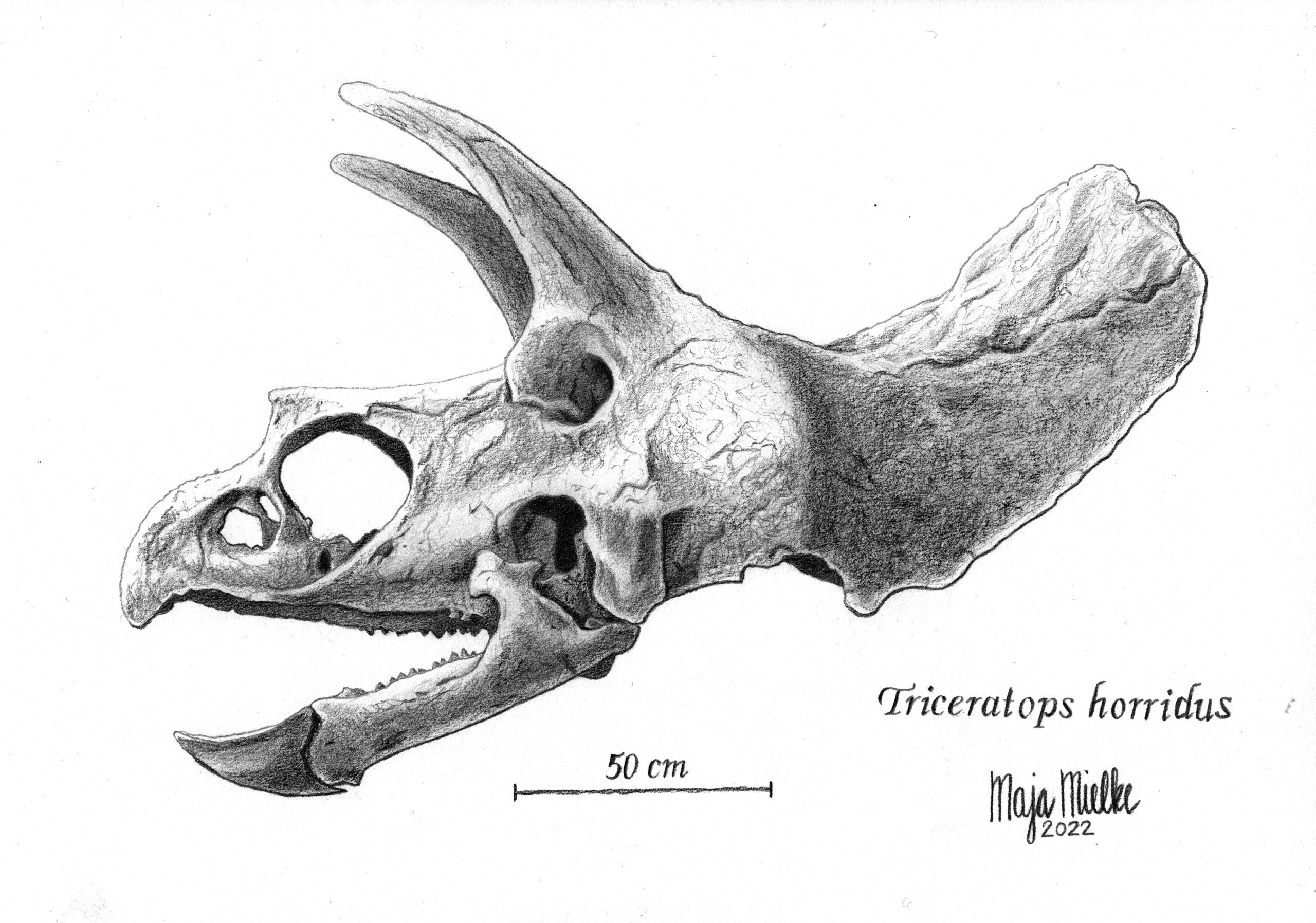
Skull of a Triceratops (pencil).Pencil is the medium I feel most comfortable with. I made this illustration after following the “Scientific Illustration” course by Oscar Sanisidro from Transmitting Science in 2022.
How do you make your art?
Most of the time, I draw from reference photos. I usually work in pencil or ink, sometimes with watercolor, and very rarely digitally. Because of my limited free time during normal working days, I often finish a drawing during multiple sessions spread over the evenings of several days. If I’m not working on an elaborate piece, I mostly just practice some basic drawing skills with quick sketches in my sketchbook.
Red fox (pen and ink and watercolor in sketchbook). This fox started as a quick sketch in my sketchbook, without the intention to ‘finalize’ it in any way. But sometimes, drawings make their own developments.
Does your art influence your science at all, or are they separate worlds?
Sometimes, it’s more the other way around: my science influences my art. I create my own scientific illustrations, e.g., for conference presentations or papers. Also, because I study birds, I particularly enjoy drawing them. But most of the time, my art is doing its own thing, just exploring the animal world and pausing for a sketch whenever inspiration hits.
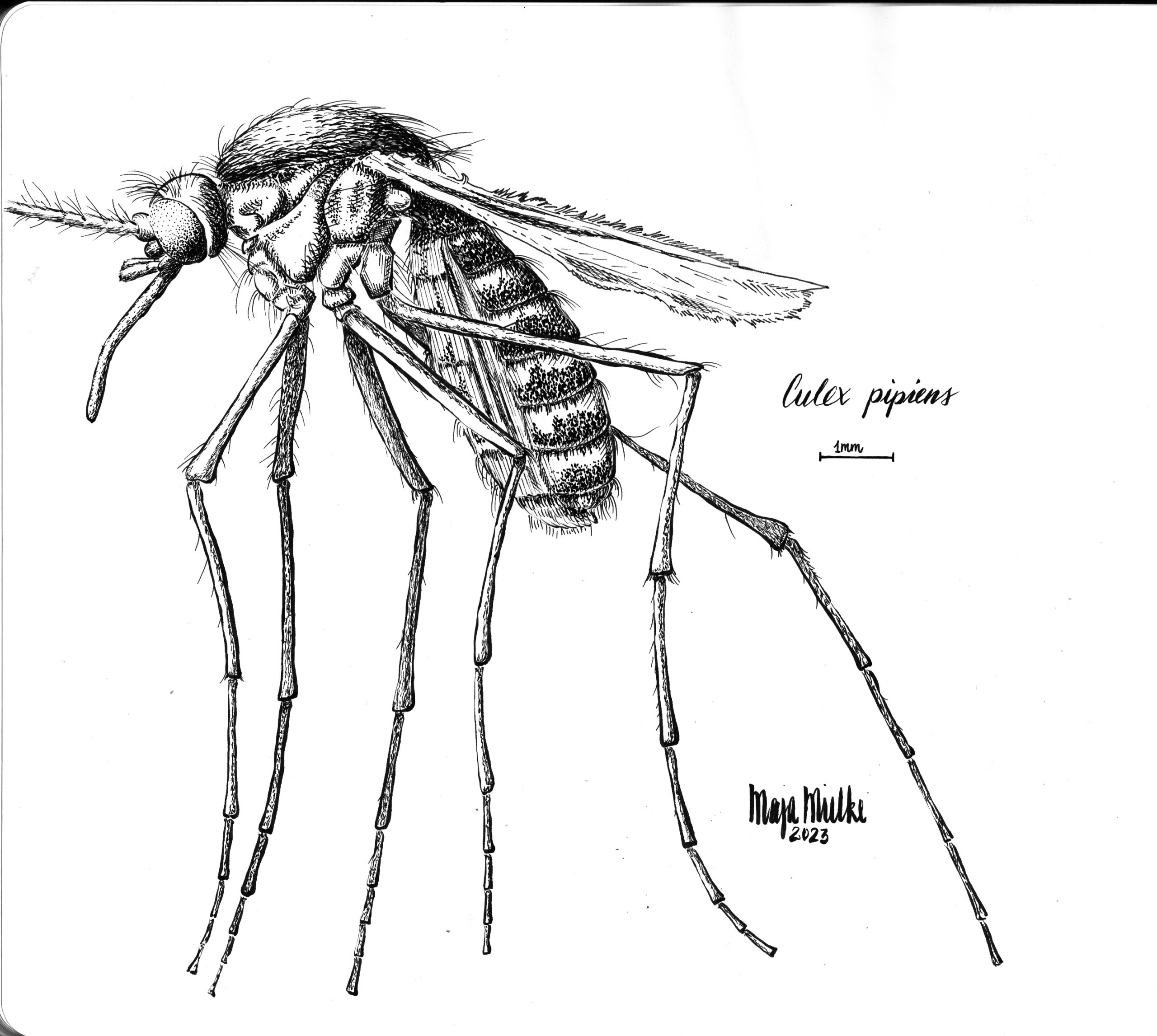
Common house mosquito (pen and ink in sketchbook). Normally, I’m more interested in vertebrates (both for my research and as my drawing subjects). But insects are just so much fun to draw. The reference photo for this drawing is from Rick Delhem’s book “Een boek vol insecten”.
What are you thinking of working on next?
I would like to further improve my pen and ink drawing skills and explore drawing with dip pens. I love the minimalistic approach of using only black ink as a medium instead of dozens of colors. Visualizing different textures like feathers, fur, and scales with only ink lines or dots is challenging but also so much fun.
Do you study Development? Complex cellular systems? Cancer? Then we’d love to have you at our Meeting “Cell Lineages across scales, space and time”!
Lineage trajectories and cellular decisions coordinate embryonic development and growth. Similarly, cellular origin and clonal trajectories are crucial elements to understanding and modulating the response to injury, disease, and therapies. This meeting will showcase cutting-edge research spanning developmental biology, regeneration, and cancer, in light of recently developed techniques to study lineage trajectories and cellular decisions with different approaches.
There’s something for everyone, whether you’re into classic lineage tracing/fate mapping, imaging-based cell tracking, genomic barcoding, or somatic mutation tracing. Come interact with experts on these techniques, covering development, regeneration, and disease, in classical and non-classical model organisms.
Registration is free – just go to the website and request an invitation:
Simon Hippenmeyer – IST, Austria Ben Simons – Cambridge University, UK Shosei Yoshida – NIBB Okazaki, Japan Elizabeth Murchison – Cambridge University, UK Sarah Bowling – Boston Children’s Hospital -USA Bushra Raj – UPenn – USA Elke Ober- FAU Erlangen Nurnberg, Germany Trevor Graham – ICR London, UK Patrick Lemaire – CRBM, Montpellier – France Colinda Scheele – IB-KU Leuven, Belgium Michael Ratz – Karolinska Institute, Sweden Michel Cayouette – IRCM Montreal, Canada Periklis Pantazis – Imperial College London, UK Ana Cvejic – University of Copenhagen/BRIC – Denmark Marketa Kaucka – MPRGL MPI – Germany Mekayla Storer – Cambridge Stem Cell Institute, UK
Vasileios R. Ouzounidis, Mattie Green, Charlotte de Ceuninck van Capelle, Clara Gebhardt, Helena Crellin, Cameron Finlayson, Bram Prevo, Dhanya K. Cheerambathur
Kristen Kurtzeborn, Vladislav Iaroshenko, Tomáš Zárybnický, Julia Koivula, Heidi Anttonen, Darren Brigdewater, Ramaswamy Krishnan, Ping Chen, Satu Kuure
Jenifer P. Suntharalingham, Ignacio del Valle, Federica Buonocore, Sinead M. McGlacken-Byrne, Tony Brooks, Olumide K. Ogunbiyi, Danielle Liptrot, Nathan Dunton, Gaganjit K Madhan, Kate Metcalfe, Lydia Nel, Abigail R. Marshall, Miho Ishida, Neil J. Sebire, Gudrun E. Moore, Berta Crespo, Nita Solanky, Gerard S. Conway, John C. Achermann
Nikola Sekulovski, Jenna C. Wettstein, Amber E. Carleton, Lauren N. Juga, Linnea E. Taniguchi, Xiaolong Ma, Sridhar Rao, Jenna K. Schmidt, Thaddeus G. Golos, Chien-Wei Lin, Kenichiro Taniguchi
Antonia Hauth, Jasper Panten, Emma Kneuss, Christel Picard, Nicolas Servant, Isabell Rall, Yuvia A. Pérez-Rico, Lena Clerquin, Nila Servaas, Laura Villacorta, Ferris Jung, Christy Luong, Howard Y. Chang, Judith B. Zaugg, Oliver Stegle, Duncan T. Odom, Agnese Loda, Edith Heard
Federica Buonocore, Jenifer P Suntharalingham, Olumide K Ogunbiyi, Aragorn Jones, Nadjeda Moreno, Paola Niola, Tony Brooks, Nita Solanky, Mehul T. Dattani, Ignacio del Valle, John C. Achermann
Shruthi Bandyadka, Diane PV Lebo, Albert Mondragon, Sandy B Serizier, Julian Kwan, Jeanne S Peterson, Alexandra Y Chasse, Victoria Jenkins, Anoush Calikyan, Anthony Ortega, Joshua D Campbell, Andrew Emili, Kimberly McCall
Sofia E. Luna, Joab Camarena, Jessica P. Hampton, Kiran R. Majeti, Carsten T. Charlesworth, Eric Soupene, Sridhar Selvaraj, Kun Jia, Vivien A. Sheehan, M. Kyle Cromer, Matthew H. Porteus
Haitham Sobhy, Marco De Rovere, Amina Ait-Ammar, Muhammad Kashif, Clementine Wallet, Fadoua Daouad, Thomas Loustau, Carine Van Lint, Christian Schwartz, Olivier Rohr
Barbara Zhao, Jacob Socha, Andrea Toth, Sharlene Fernandes, Helen Warheit-Niemi, Brandy Ruff, Gurgit K. Khurana Hershey, Kelli L. VanDussen, Daniel Swarr, William J. Zacharias
Gerrald A. Lodewijk, Sayaka Kozuki, Clara Han, Benjamin R. Topacio, Abolfazl Zargari, Seungho Lee, Gavin Knight, Randolph Ashton, Lei S. Qi, S. Ali Shariati
Paula A. Balestrini, Abdelbaki Ahmed, McCarthy Afshan, Liani Devito, Claire E. Senner, Alice E. Chen, Prabhakaran Munusamy, Paul Blakeley, Kay Elder, Phil Snell, Leila Christie, Paul Serhal, Rabi A. Odia, Mahesh Sangrithi, Kathy K. Niakan, Norah M.E. Fogarty
Tania Hübscher, L. Francisco Lorenzo-Martín, Thomas Barthlott, Lucie Tillard, Jakob J. Langer, Paul Rouse, C. Clare Blackburn, Georg Holländer, Matthias P. Lutolf
Abdulvasey Mohammed, Benjamin D. Solomon, Priscila F. Slepicka, Kelsea M. Hubka, Hanh Dan Nguyen, Wenqing Wang, Martin Arreola, Michael G. Chavez, Christine Y. Yeh, Doo Kyung Kim, Virginia Winn, Casey A. Gifford, Veronika Kedlian, Jong-Eun Park, Georg A. Hollander, Vittorio Sebastiano, Purvesh Khatri, Sarah A. Teichmann, Andrew J. Gentles, Katja G. Weinacht
Lulu Huang, Jochem H. Bernink, Amir Giladi, Daniel Krueger, Gijs J.F. van Son, Maarten H. Geurts, Georg Busslinger, Lin Lin, Maurice Zandvliet, Peter J. Peters, Carmen Lopez-Iglesias, Christianne J. Buskens, Willem A. Bemelman, Harry Begthel, Hans Clevers
Onur Eskiocak, Saria Chowdhury, Vyom Shah, Emmanuella Nnuji-John, Charlie Chung, Jacob A. Boyer, Alexander S. Harris, Jill Habel, Michel Sadelain, Semir Beyaz, Corina Amor
Augusto Ortega Granillo, Daniel Zamora, Robert R. Schnittker, Allison R. Scott, Jonathon Russell, Carolyn E. Brewster, Eric J. Ross, Daniel A. Acheampong, Kevin Ferro, Jason A. Morrison, Boris Y. Rubinstein, Anoja G. Perera, Wei Wang, Alejandro Sánchez Alvarado
Lucia Leitner, Martina Schultheis, Franziska Hofstetter, Claudia Rudolf, Valeria Kizner, Kerstin Fiedler, Marie-Therese Konrad, Julia Höbaus, Marco Genini, Julia Kober, Elisabeth Ableitner, Teresa Gmaschitz, Diana Walder, Georg Weitzer
Qian Ma, Sijia Liu, Sara Raggi, Siamsa M. Doyle, Barbora Pařízková, Deepak Kumar Barange, Edward G. Wilkinson, Isidro Crespo Garcia, Joakim Bygdell, Gunnar Wingsle, Dirk Roeland Boer, Lucia C. Strader, Fredrik Almqvist, Ondřej Novák, Stéphanie Robert
Xiaosa Xu, Michael Passalacqua, Brian Rice, Edgar Demesa-Arevalo, Mikiko Kojima, Yumiko Takebayashi, Benjamin Harris, Hitoshi Sakakibara, Andrea Gallavotti, Jesse Gillis, David Jackson
Amir Fallahshahroudi, Leticia Rodríguez-Montes, Sara Yousefi Taemeh, Nils Trost, Memo Tellez Jr., Maeve Ballantyne, Alewo Idoko-Akoh, Lorna Taylor, Adrian Sherman, Enrico Sorato, Martin Johnsson, Margarida Cardoso Moreira, Mike J. McGrew, Henrik Kaessmann
Aurélie Hintermann, Christopher Chase Bolt, M. Brent Hawkins, Guillaume Valentin, Lucille Lopez-Delisle, Sandra Gitto, Paula Barrera Gómez, Bénédicte Mascrez, Thomas A. Mansour, Tetsuya Nakamura, Matthew P. Harris, Neil H. Shubin, Denis Duboule
Steven L. Brody, Jiehong Pan, Tao Huang, Jian Xu, Huihui Xu, Jeffrey Koenitizer, Steven K. Brennan, Rashmi Nanjundappa, Thomas G. Saba, Andrew Berical, Finn J. Hawkins, Xiangli Wang, Rui Zhang, Moe R. Mahjoub, Amjad Horani, Susan K. Dutcher
Pooja Popli, Arin K. Oestreich, Vineet K. Maurya, Marina N. Rowen, Ramya Masand, Michael J. Holtzman, Yong Zhang, John Lydon, Shizuo Akira, Kelle H. Moley, Ramakrishna Kommagani
Shruthi Bandyadka, Diane PV Lebo, Albert Mondragon, Sandy B Serizier, Julian Kwan, Jeanne S Peterson, Alexandra Y Chasse, Victoria Jenkins, Anoush Calikyan, Anthony Ortega, Joshua D Campbell, Andrew Emili, Kimberly McCall
Francisco J. Romero-Expósito, Almudena Moreno-Rivero, Marta Muñoz-Barrera, Sabas García-Sánchez, Fernando Rodríguez-Peris, Nicola Gritti, Francesca Sartor, Martha Merrow, Jeroen S. van Zon, Alejandro Mata-Cabana, María Olmedo
Enikő Lázár, Raphaël Mauron, Žaneta Andrusivová, Julia Foyer, Ludvig Larsson, Nick Shakari, Sergio Marco Salas, Sanem Sariyar, Jan N. Hansen, Marco Vicari, Paulo Czarnewski, Emelie Braun, Xiaofei Li, Olaf Bergmann, Christer Sylvén, Emma Lundberg, Sten Linnarsson, Mats Nilsson, Erik Sundström, Igor Adameyko, Joakim Lundeberg
Lara Feulner, Florian Wünnemann, Jenna Liang, Philipp Hofmann, Marc-Phillip Hitz, Denis Schapiro, Severine Leclerc, Patrick Piet van Vliet, Gregor Andelfinger
Andreana Gomez, Sergio Gonzalez, Ashwini Oke, Jiayu Luo, Johnny B. Duong, Raymond M. Esquerra, Thomas Zimmerman, Sara Capponi, Jennifer C. Fung, Todd G. Nystul
We aim to understand how human developmental cell fate decisions are controlled and exploit this knowledge for regenerative medicine and disease modelling applications. We are focusing particularly on defining how embryonic cells acquire an anteroposterior (A-P) axial identity and its effect on disease vulnerability and developmental potency/plasticity. To address our questions, we employ human pluripotent stem cell (hPSCs) differentiation as an in vitro model of early human embryonic development and utilise protocols we have recently established toward the production of human spinal cord progenitors and neural crest cells that correspond to distinct axial levels.
Current projects in our lab include:
1) Understanding how various cellular components of the trunk (e.g. motor neurons, paraxial mesoderm, trunk neural crest) are derived from multipotent axial progenitors.
2) Defining the molecular/signalling basis of A-P axial identity acquisition/maintenance in distinct neural cell types.
3) Deciphering the molecular/signalling logic of cell fate decisions in vagal neural crest derivatives focusing on the enteric nervous system and the treatment of enteric neuropathies such as Hirschsprung disease.
4) Determining how abnormal trunk neural crest development is linked to the initiation of neuroblastoma, the most common extra-cranial solid tumour of childhood.
Lab roll call
James Birch: PhD student, studying the links between cell competition and neuroblastoma initiation.
Katy Boswell: PhD student, investigating the effect of chromosome 17q gain/MYCN overactivation on the enhancer landscape of hPSC-derived trunk neural crest cells.
Fay Cooper: Postdoc, studying determinants of axial identity in spinal cord progenitors and neural crest cells. She has also been involved in the development of a hPSC-based cell therapy for treating Hirschsprung disease. More recently, Fay has been examining the effect of DNA damage on trunk neural crest differentiation in hPSCs harbouring various neuroblastoma-associated aberrations.
Grace Gilbert: Joint PhD student (primary supervisor: Dr Dan Bose), dissecting the effect of Rubinstein Taybi Syndrome disease mutations on the epigenetics of neural crest/neuronal development.
Sude Uyulgan: PhD student, working on the study of axial identity acquisition by hPSC-derived lateral plate mesoderm and limb progenitors.
Favourite technique, and why?
Anestis: Immunostaining/fluorescence microscopy analysis of neuronal projections simply because it usually produces images of immense hypnotic beauty.
Apart from your own research, what are you most excited about in developmental and stem cell biology?
Anestis: I am very excited about the increasing number of complementary 2 -and 3-D hPSC differentiation models, which together with emerging cutting-edge technologies in microscopy, sequencing and bioengineering offer for the first-time unique opportunities for controlling and understanding human developmental processes. They can also facilitate examining older classic embryology experiments/ concepts under a new lens. I am very keen on using these approaches in the near future to address specifically our group’s questions.
What is the best thing about where you work?
Anestis: The University of Sheffield hosts a vibrant developmental/stem cell biology community, and I have met some fantastic colleagues, collaborators and friends here.
Sude: Supportive and friendly atmosphere, exciting research, and most importantly having Anestis as a supervisor!
James: The University of Sheffield is a great place to work. We have regular fascinating seminars and the university has many support groups from research on aging to data analysis (Sheffield R Users Group) to support you on your research journey. The university really cares what we think, I’ve seen management respond to student and staff surveys and to really be willing to put funding behind making changes where needed. I think all these contribute to making Sheffield an exciting and supportive working environment.
Fay: The people…. we have a really great dynamic in the lab at the moment which makes it a really enjoyable place to work.
What’s there to do outside of the lab?
Anestis: Sheffield is home to lots of pubs/breweries (it is currently the real ale capital of the world), restaurants, cinemas and concert venues so there are plenty of options. It is also close to the Peak District so stunning nature/landscapes are literally around the corner.
Sude: Hiking in the Peak District, great pie and good pubs.
James: Sheffield is a really vibrant and active city with loads to do. If you like music and street food we have loads of great music venues, pubs, clubs and food stalls all around the city. We’re also just a 20 minute drive from the peaks. One of my favourite hiking areas is the Edale valley, easily accessible by car and train. There are several great hiking routes and some of the highest peaks in the peak district, as well as a cozy pub (The Old Nags Head) for a drink and a great meal after a good walk.
Fay: Sheffield is the outdoor city… it is so close to the peak district which has some amazing walking, running and swimming spots. I spend as much time as I can enjoying this!
Browse through other ‘Lab meeting’ posts featuring developmental and stem cell biology labs around the world.
Daniel Wehner is Group Leader at the Max Planck Institute for the Science of Light and the Max-Planck-Zentrum für Physik und Medizin in Erlangen (Germany). At the 2024 joint German and Dutch developmental biology societies meeting in Osnabrück, Daniel was awarded the GfE Hilde Mangold Award for young investigators. After his award lecture at the conference, we chatted about his career background, his transition to becoming a PI, and his research into spinal cord regeneration in zebrafish.
(Image credit: Stephan Spangenberg)
Congratulations on winning the GfE Hilde Mangold Award! What does this award mean to you?
Receiving this award is of tremendous significance to me because it recognizes the efforts and contributions of our research team to the field. The German Society for Developmental Biology is my home turf, so I am deeply honoured to be recognized by this society for our work.
You received the GfE PhD prize back in 2015 as well!
Yes, this makes getting this Hilde Mangold Award even more impactful, as it validates that we are on the right path and encourages me to persist in my research direction. I believe our research has always been somewhat unconventional in that we never opt for the easiest route to tackle scientific challenges. For instance, our lab combines biochemical and physical methodologies to investigate spinal cord regeneration. It’s truly gratifying to see the efforts of our research group acknowledged. I extend my heartfelt thanks to all the members of my group and my past and present mentors who have supported us along the way.
Let’s go back to the beginning, when did you first become interested in science?
I have always had a keen interest in biology and in understanding the fundamental principles of life. However, it was during my final two years of high school in the biology course that my passion truly ignited. That was when I was introduced to the basic principles of cell biology, and I became captivated by the intricacies and sophistication that enable life. I went on to study (medical) biotechnology, yet I found myself unsatisfied with merely learning about established knowledge. The decision to pursue a PhD stemmed from my inherent drive to discover something new.
What did you work on during your PhD?
I completed my PhD under the supervision of Gilbert Weidinger, initially at the Technische Universität Dresden and later at Ulm University. Initially, my research focused on identifying new modulators of the Wnt pathway [1] [2]. Subsequently, I shifted my focus to investigating how Wnt signaling regulates fin regeneration. We adapted the TetON system to examine cell type-specific functions of Wnt signaling in a complex regenerating appendage [3]. This technological advancement led to the concept of Wnt pathway-controlled signaling centers that coordinate regeneration through secondary signals. [4] [5]. In another paper published in Development, we provided evidence that Notch signalling coordinates the proliferation and differentiation of progenitor cells — key events in tissue regeneration [6].
Then you moved to Edinburgh for your postdoc at the Becker lab. What did you work on?
Catherina and Thomas Becker have done pioneering work on spinal cord regeneration in zebrafish. It was an excellent synergy, bringing together the TetON system and their remarkable model system for central nervous system regeneration. It was during this time that we established the principle that after spinal cord injury, fibroblasts modulate the lesion site by depositing a complex extracellular matrix (ECM), but in strong contrast to mammals, in zebrafish this environment promotes regeneration [7].
Is that when you started to find your niche for your own research group?
Yes, I was intrigued by the interspecies difference in fibroblast behaviour because, in mammals, fibroblasts are typically viewed as inhibiting the regeneration of the central nervous system. For instance, research conducted by the Frisén and Göritz labs demonstrated that PDGFRβ-positive fibroblasts of perivascular origin contribute to the formation of inhibitory central nervous system scars [8] [9]. Therefore, the natural question to ask was: do these cells behave differently in response to spinal cord injury in zebrafish? To address this, we developed numerous genetic models to lineage-trace and cell type-specifically interfere with pdgfrb+ cell recruitment, or spatially-confined optogenetic depletion of pdgfrb+ cells. Our findings revealed that in zebrafish, pdgfrb+ fibroblasts are required for regeneration. We discovered that these fibroblasts possess the ability to downregulate specific inhibitory ECM molecules and upregulate regeneration-promoting matrix genes, facilitating axon regeneration. We further demonstrated that the unique biochemical composition of fibroblast-derived injury ECM in central nervous system lesions defines regenerative success, versus inhibitory scar formation in zebrafish and mammals. These findings resulted in the publication of our lab’s first two major papers [10] [11].
How was your experience transitioning to becoming a PI?
I must admit that the transition was not easy. There were moments when I considered leaving academia due to the lack of prospects. This is a challenge many colleagues face, particularly women in the field. As a father of two, I understand the difficulties and challenges of navigating a postdoc while caring for children, and I know firsthand the additional challenges young parents encounter. I consider myself very fortunate that things have ultimately worked out for me.
Following my postdoc in Edinburgh, I joined the Center for Regenerative Therapies at Technische Universität Dresden, working in Michael Brand’s lab with support from a return stipend from the German Research Foundation. Although I applied for an Emmy Noether Fellowship (similar to an ERC starting grant) and didn’t make the final cut, it was during this time that I crossed paths with Jochen Guck, a renowned researcher in tissue mechanics. We discovered excellent synergies by combining physics methodologies with molecular biology tools to investigate spinal cord regeneration in zebrafish. This led to an exciting opportunity to lead my own independent research group at the Max Planck Institute for the Science of Light.
Can you briefly talk about what your lab is working on?
My current research addresses the fundamental questions: i) How can regeneration-permissive lesion environments be established after central nervous system injury? ii) How can central nervous system axon regeneration be promoted in regeneration-limiting environments?
To achieve these goals, we are pursuing several approaches. i) We are investigating the intricate cell-cell interactions that dictate the biochemical and mechanical properties of the regeneration-permissive microenvironment in zebrafish, including the regulation of axon regeneration-permissive injury ECM. ii) Utilizing zebrafish as an in vivo platform, we are dissecting the regeneration-modulating properties of specific components of human scar tissue, along with elucidating their underlying mechanisms. iii) We are also in the process of developing humanized zebrafish models to explore novel strategies for spinal cord repair.
Do you have any advice for people thinking of starting their own lab?
Science thrives on passion and determination, existing within its own realm. Avoid comparing yourself to those outside of this world. Instead, dare to think innovatively and seek out environments that foster support, enabling you to embrace risks and confront challenges.
Speaking of supportive environments, how important do you think mentorship is in navigating an academic career?
A supportive mentor can profoundly influence career development. Navigating the academic path is challenging and fraught with obstacles. Having a great mentor who offers guidance on avoiding these pitfalls can be invaluable, sparing you from unnecessary setbacks.
I count myself fortunate to have benefitted from excellent mentorship. For those seeking PhD or postdoc positions, I recommend considering joining a lab led by a junior PI. Working with a junior PI provides valuable insight into the process of establishing a lab. Junior PIs often prioritize mentorship as it is a mutual responsibility.
My PhD supervisor, Gilbert, was a junior PI at the time. He taught me essential techniques such as microinjections, then entrusted me with the freedom to explore. This supportive environment was echoed during my time at Becker’s lab in Edinburgh, where guidance was readily available, yet I was encouraged to pursue independent research.
What’s your favourite technique? And what are you most excited about in your area?
My preferred technique is in situ hybridization—it’s remarkably reliable! Nevertheless, the method that currently captivates me the most is Brillouin microscopy. This non-invasive, label-free, all-optical approach enables the measurement of viscoelastic properties in vivo using light. Access to Brillouin microscopes is scarce, limited to only a handful of locations worldwide. Having the chance to employ this state-of-the-art technique at the Max Planck Institute in Erlangen to investigate the role of tissue mechanics in spinal cord regeneration is a true privilege. Achieving the initial results that contributed to our latest paper [11] was an overwhelming experience.
Finally, let’s get outside of the lab – what do you like to do in your free time?
Back then, I was deeply immersed in music, playing bass guitar in a band, and engaging in trekking, rock climbing, and mountaineering. However, with the arrival of my two daughters and the commencement of a new lab, I find myself with limited time for hobbies. Nonetheless, I consider science itself a passion—a hobby in its own right!
(Image credit: Stephan Spangenberg)
References
[1] Kagermeier-Schenk B, Wehner D, Ozhan-Kizil G, et al. Waif1/5T4 inhibits Wnt/β-catenin signaling and activates noncanonical Wnt pathways by modifying LRP6 subcellular localization. Dev Cell. 2011;21(6):1129-1143. doi:10.1016/j.devcel.2011.10.015
[2] Özhan G, Sezgin E, Wehner D, et al. Lypd6 enhances Wnt/β-catenin signaling by promoting Lrp6 phosphorylation in raft plasma membrane domains. Dev Cell. 2013;26(4):331-345. doi:10.1016/j.devcel.2013.07.020
[3] Wehner, D., Jahn, C., Weidinger, G. Use of the TetON System to Study Molecular Mechanisms of Zebrafish Regeneration. J. Vis. Exp.(100), e52756, doi:10.3791/52756 (2015)
[4] Wehner D, Cizelsky W, Vasudevaro MD, et al. Wnt/β-catenin signaling defines organizing centers that orchestrate growth and differentiation of the regenerating zebrafish caudal fin [published correction appears in Cell Rep. 2014 Feb 27;6(4):777-8]. Cell Rep. 2014;6(3):467-481. doi:10.1016/j.celrep.2013.12.036
[5] Wehner D, Weidinger G. Signaling networks organizing regenerative growth of the zebrafish fin. Trends Genet. 2015;31(6):336-343. doi:10.1016/j.tig.2015.03.012
[6] Bartholomäus Grotek, Daniel Wehner, Gilbert Weidinger; Notch signaling coordinates cellular proliferation with differentiation during zebrafish fin regeneration. Development 1 April 2013; 140 (7): 1412–1423. doi: 10.1242/dev.087452
[7] Wehner D, Tsarouchas TM, Michael A, et al. Wnt signaling controls pro-regenerative Collagen XII in functional spinal cord regeneration in zebrafish. Nat Commun. 2017;8(1):126. Published 2017 Jul 25. doi:10.1038/s41467-017-00143-0
[8] Dias DO, Kim H, Holl D, et al. Reducing Pericyte-Derived Scarring Promotes Recovery after Spinal Cord Injury. Cell. 2018;173(1):153-165.e22. doi:10.1016/j.cell.2018.02.004
[9] Göritz C, Dias DO, Tomilin N, Barbacid M, Shupliakov O, Frisén J. A pericyte origin of spinal cord scar tissue. Science. 2011;333(6039):238-242. doi:10.1126/science.1203165
[10] Tsata V, Möllmert S, Schweitzer C, et al. A switch in pdgfrb+ cell-derived ECM composition prevents inhibitory scarring and promotes axon regeneration in the zebrafish spinal cord. Dev Cell. 2021;56(4):509-524.e9. doi:10.1016/j.devcel.2020.12.009
[11] Kolb, J., Tsata, V., John, N. et al. Small leucine-rich proteoglycans inhibit CNS regeneration by modifying the structural and mechanical properties of the lesion environment. Nat Commun 14, 6814 (2023). https://doi.org/10.1038/s41467-023-42339-7
This is an Editorial in Volume 151, Issue 6 of Development, written by Alex Eve and Oliver Hobert.
This year, 2024, marks the 40th year of the discovery of the homeobox, a landmark discovery that fundamentally impacted multiple fields, ranging from genetics and genomics to developmental biology, neuroscience and evolution. The application of molecular biology techniques to analyze mutant animals, beginning in the 1980s, allowed two laboratories on two different continents to independently find that classic Drosophila mutants with characteristic developmental patterning defects carried sequence alterations in specific genetic loci which shared an ∼180 nucleotide sequence called the ‘homeobox’. This name arose from the intriguing phenotypic alterations observed in homeobox gene mutants, namely the transformation of the identity of some body part into that of another body part. Such identity transformations had long been known to exist in animals and plants, and had been dubbed ‘homeotic transformations’ by William Bateson, one of the fathers of modern-day genetics. The first Drosophila mutants with such a transformation were first identified by Calvin Bridges in Thomas Hunt Morgan’s legendary Fly Room at Columbia University, and further characterized by Ed Lewis at Caltech, whose work on homeobox genes earned him the Nobel Prize in 1995. After the molecular identification of homeobox genes in 1984 by Michael Levine and Bill McGinnis in Walter Gehring’s lab in Switzerland (McGinnis et al., 1984) and Matt Scott and Amy Weiner in Thomas Kaufman’s lab in the USA (Scott and Weiner, 1984), it quickly became clear that the homeobox encoded a DNA-binding domain, the so-called homeodomain. The monumental impact of defining the homeobox genes did not merely lie in understanding how specific developmental patterning events are genetically specified in Drosophila, but it rather lay in the realization that this domain is conserved throughout the animal kingdom and, eventually, that not only their sequence but also their function in developmental patterning is conserved. In fact, homeobox genes exist in unicellular organisms, but their number exploded with the advent of multicellularity, indicating that they have served as key drivers in the generation of the astounding complexity of animal body plans and cell types.
Another striking realization that came from studies across the animal kingdom was that homeobox genes form a part of a large family of genes encoding transcription factors, only a subset of which control developmental patterning along the anterior-posterior axis of a developing embryo (the genomically clustered HOX genes). Meanwhile, a much larger set of homeobox genes are involved in controlling the development of several other cell types, most notably and perhaps most predominantly so, the nervous system. Such neuronal functions of homeobox genes are deeply conserved in animals.
To celebrate this anniversary, Development has commissioned a series of review-type articles from leaders in the field demonstrating the impact that the homeobox discovery has had on different disciplines. This series, published throughout 2024, begins in this issue with two first-hand Perspectives that reflect on ‘perhaps the most exciting aspect of the homeotic gene story’ (Heasman et al., 1985): the first evidence that the Drosophila homeobox sequence is conserved in other animal species.
In the first article, Matthew Scott lays out the historical context for the discovery following studies of the Drosophila homeotic genes and describes from personal experience the work from Indiana University, USA, in identifying a conserved homeobox sequence (Scott, 2024). In its companion piece, Bill McGinnis and Michael Levine reminisce on their own ‘eureka moment’ from their studies in Basel, Switzerland, showing that the Drosophila homeobox sequence is conserved in other animals (McGinnis and Levine, 2024). Together, both Perspectives provide a fascinating and entertaining read, capturing the excitement of discovery and the maturation of competition into lifelong friendships.
To read the series as it grows, please visit the dedicated subject collection here: https://journals.biologists.com/dev/collection/10249/40-years-of-the-homeobox. To date, Development has published almost 5000 articles on homeobox genes and, of course, we welcome your future submissions detailing how this remarkable family functions in development, stem cells and regeneration.
We are located in Lund, a small city in Southern Sweden. The University was founded in 1666, and consists of many old beautiful buildings. The lab is affiliated with the Division of Pediatrics, and we’re part of the Lund Stem Cell Center.
We are intrigued by how we can utilize the setting of normal embryonic development to understand tumor initiation. Our focus lies primarily on how cancer forms neuroblastoma and paraganglioma develop during embryogenesis. These cancers arise in the adrenal gland and along (para)sympathetic ganglia, and originate from cells of the trunk neural crest stem cell population. Neuroblastoma is the most common tumor form in infants. It is believed to arise due to developmental defects during embryogenesis, but the precise origin and cause of initiation is unknown. Paragangliomas are on the other hand adult slow growing tumors, but also these are believed to be primed during development of the neural crest. We mainly use human patient-derived cells and chick embryos to dissect the role of neural crest specific genes in cancer initiation and progression. We develop techniques where we can analyze full-scale organogenesis and tumor formation in terms of the genomic, proteomic, and morphological landscape following genetic insults in the trunk neural crest stem cell population during embryonic development.
Lab roll call
Sinan Karakaya is a postdoc, exploring the role of a dysregulated hypoxic response in both normal development and the progression of tumors, specifically focusing on paraganglioma. His research investigates the role of HIF-2α in tumor initiation, development, and aggressiveness. Sinan is originally from Turkey, and did his PhD in Germany.
Perrine Burdeyron is a postdoc working on developing a model to study neuroblastoma by using human pluripotent stem cells with chick embryo as a host. This model allows us to study different genes involved in this childhood cancer. Perrine is originally from France.
Tom Gregor is a postdoc working on the role of HIF-2α during neural crest stem cell-derived childhood cancer progression. Tom is originally from Czech Republic.
Marina Mazariegos is working as a lab engineer, and arrived to the lab six weeks ago. She is involved in several projects with different people, as well as general management and organization of the lab. Marina is originally from Spain.
Niklas Engström is a lab engineer involved in a number of projects, as well as in maintaining the structure of the lab.
Elina Fredlund is a final year PhD candidate working to understand the initiation of neuroblastoma with main focus on an early development protein that has an important role in embryogenesis and a suppressive function in neuroblastoma.
Stina Andersson is a second year PhD student, and her work focuses on how different types of neuroblastoma cells respond to various cues from the embryonic microenvironment.
Zana Stirn is a master student. She is working together with our postdoc Perrine on the project to develop a human-chick chimera model, that will be used to manipulate different genes and pathways in neuroblastoma and consequently to get a better insight into the development and origin of this cancer. Zana is originally from Slovenia.
Pleun Jornick is a master student, fulfilling the last part of her master internship here. She studies the mechanisms of a tumor-suppressor gene in neuroblastoma. Pleun is originally from The Netherlands.
Özgür Rubar Altin is a medical doctor at the beginning of his career, and he is doing an internship in the lab. He is working on investigating the role of HIF-2α in chicken embryos and different tumor cells. Özgüris originally from Turkey.
Inés Sanz is a master student, investigating the role of how embryonic microenvironmental factors affect neuroblastoma cell behavior. Inés is originally from Spain
Favourite technique, and why?
Sofie Mohlin: There are so many techniques that we can use, each with its own advantages and beauty. But I’m going to go with a ‘simple’ on: microinjections and electroporation in chick embryos. It is such a basic technique that’s been around for ages, yet it’s the basis for so many experiments and essential knowledge we have on embryogenesis today. Unfortunately, I don’t spend a lot of time on hands-on lab work anymore, but when I do it’s always by the microscope doing injections. While it in one way requires focus and concentration, I tend to go into a meditative state where I can let go of stress and just enjoy the beauty of the embryos, one after one after one…
Apart from your own research, what are you most excited about in developmental and stem cell biology?
Sofie Mohlin: I think the process of (neural crest) stem cell lineage commitment is so delicate and exciting. With the recent in-depth sequencing techniques, we’ve come a long way in understanding the genetics behind, and as a result of, lineage commitment. However, there is so much left to understand about the biology surrounding lineage commitment.
How do you approach managing your group and all the different tasks required in your job?
Sofie Mohlin: My philosophy is that people are given full independence, but it comes with responsibility. In the lab we drive the projects forward together, but each individual project is one person’s pet project (or, someone’s baby, which is my favorite way of expressing it). However, I make sure I’m always available, as often as possible physically in the lab, otherwise being quick in answering questions via email/chat functions. We also have time-fixed regular lab meetings and one-on-one meetings with PhD students. Planning and prioritization are key, but to be honest it’s not always easy. With deadlines, grant applications and academic commitments, I often feel that time for actual science is sparse. I try to be open to the group when very intense periods occur so that they know why I might be more stressed and less available for a (hopefully short) while. I’m also working on being able to say no to things. There are so many things to do, and many of these are fun tasks. But also, with a limited number of hours per day, being able to prioritize and say no is crucial. At the end of the day, the time when I’m the happiest at work is after meetings with lab members, where we have discussed cool new data, experiments, and future plans, leaving with the feeling of exhilaration. This is an activity that I would never skip!
What is the best thing about where you work?
Sofie Mohlin: My group of course! I really enjoy discussing and doing science, as well as chatting about everyday life, with everyone, be it in the corridor, in the lab, in one-on-one meetings or lab meetings. I also work close with other great PIs, providing me with a fantastic peer community and surrounding.
Sinan Karakaya: The best part of working in our research lab is the collaborative environment fostered by our diverse team. Each member brings unique perspectives and expertise, creating an atmosphere conducive to innovative thinking and problem-solving. Additionally, the supportive and encouraging nature of Sofie and colleagues nurture professional growth and fosters a sense of camaraderie, making it an enriching and fulfilling experience to be part of our research endeavors.
Perrine Burdeyron: The best thing is the positive environment in the lab.
Marina Mazariegos: Definitely the work environment created by great people working for the good of the whole lab and not for themselves. And also, the prospect of achieving goals to contribute to progress in clinical research.
Elina Fredlund: The best things are my colleagues and the abundant opportunities available. Guidance is readily accessible upon request, as people are very friendly and supportive.
Stina Andersson: I love learning new things every day, and I really appreciate the collaborative atmosphere in our research group.
Zana Stirn: The environment! I really enjoy the dynamic of the group. We have many get togethers, workshops, potlucks, fikas. All of this contributes to an amazing environment, which makes coming to work even more enjoyable.
Pleun Jornick: The ambiance in the lab is amazing. It feels you can always go to anyone with questions and that your opinion is really appreciated. This makes me really motivated to give my all.
Özgür Rubar Altin: Excellent colleagues and scientific working environment.
What’s there to do outside of the lab?
Sofie Mohlin: Lund is a very picturesque city with lots of beautiful old buildings and surroundings, so just walking around town is nice. Ten minutes on the train then takes you to Malmö, a wonderful city with lots of culture, sightseeing, parks, food, and wine bars!
Sinan Karakaya: Outside of the lab, there are plenty of opportunities to spend your time indoors or outdoors. Besides being a scenic, historical town, Lund also offers variety of indoor and outdoor activities including museums, sports halls and trails for trekking and biking.
Perrine Burdeyron: During the winter you can enjoy ice skating and snow activities and during the summer you can enjoy the long days to go for hiking and visit all the national parks in the south. If you prefer the city, we are close to Copenhagen, the Danish capital.
Marina Mazariegos: It´s a small city but you can find a lot of activities and there´re a lot of nice places to go to relax with friends.
Elina Fredlund: Crossing the bridge, Copenhagen is just a 40 min train ride away. Söderåsen is a beautiful national forest that’s worth a visit during any season.
Stina Andersson: I enjoy spending my free time in nature or at dinner with friends and family.
Zana Stirn: Lund is like a big student campus, so the student life is very much alive. You can enjoy different cafes, bars, parks and a beautiful botanical garden. On the weekend, you can make a trip to Lomma beach or one of many natural parks in the area. For some bigger city vibes, Malmö and Copenhagen are only a short train ride away.
Pleun Jornick: As an international student in Sweden there is a lot to discover, but the tradition I cherish the most here is “fika”. A coffee break with a sweet pastry a few times a week is a celebration. There are a lot of different nice cafés in Lund, so the challenge is to try them all.
Özgür Rubar Altin: Lund has incredible nature that you can explore by bike. There is a nice football stadium where you can go to watch Lunds BK.
Browse through other ‘Lab meeting’ posts featuring developmental and stem cell biology labs around the world.
 (No Ratings Yet)
(No Ratings Yet)





 (3 votes)
(3 votes)