Everyone is different, but we all start as a fertilised egg – we’re made the same way.
Scientists in the Wellcome-funded Human Developmental Biology Initiative are trying to understand more about how the human body is formed from fertilisation to birth. Eventually, this understanding will enable better treatments for many conditions such as heart defects, spina bifida, childhood cancers and infertility. However, human developmental biology is a difficult type of research to talk about – it involves working with human embryos and fetal tissues and the health implications aren’t exactly known yet.
In our new podcast, ‘Made the Same Way’, young artists collaborate with scientists, embracing challenging conversations about this research. In each episode, an artist creates an original song or poem which echos their feelings and stimulates curiosity about research in human developmental biology. We produced the podcast in collaboration with Reform Radio.
In the first episode, scientist Emma Rawlins meets singer-songwriter Karis Jade to discuss the science of human development. They chat about the basics; what is human developmental biology? Why is it worth studying? And what will it be used for in the future? Throughout the episode, the pair will write and record an original piece of music inspired by their meeting, exploring science in a brand new way.
You can watch a short documentary about the making of the our first episode here.
Please subscribe and listen on Apple podcasts, Spotify, or wherever you get your podcasts. If you enjoy the podcast, please rate and review us on Apple podcasts to help others find us!
I am Joyce Yu, the new Community Manager of the Node. Before joining The Company of Biologists, I was doing science communications at Alzheimer’s Research UK, a charity for dementia research. Prior to that, I did my PhD and a short postdoc in the lab of Jean-Paul Vincent at the Francis Crick Institute, where I studied the role of Wnt signalling in growth and planar cell polarity of fruit flies.
During my PhD, I found the Node to be a useful resource and I enjoyed reading a wide variety of posts on the Node. Fast forward several years, I am now very excited to be joining the Node and contributing to this invaluable community that I once benefited from immensely.
Helen and the previous Community Managers have done an amazing job producing content and engaging with the developmental and stem cell research community. I hope to build on their work and continue working on existing features, while testing out some new ideas and initiatives.
I would like to remind you that once you are registered with the Node, you can immediately start posting on the site. Anyone is welcome to share blog posts, job adverts, and events notice within the community. If you have any ideas or questions about the Node, don’t hesitate to get in touch at thenode@biologists.com or drop me an email.
I look forward to getting to know all of you here at the Node!
If we’re going to do science communication or science engagement well, we’ve got to do emotions as well as facts. So I think pop culture’s really helpful there.
Dr Jonathan Roberts
In the latest episode of the Genetics Unzipped podcast, we’re taking a leftfield look at public communication of genetics, exploring how the science of DNA turns up in popular culture from comics and music to cakes, with Great British Bake Off winner Syabira Yusoff.
We recently announced that we will be working with three newly appointed the Node correspondents, who will be helping us to develop and write content for the Node in 2023. We caught up with each of them to chat about their research backgrounds and the topics that they’re excited to write about over the course of the coming year.
Working alongside Brent Foster, we have Alexandra Bisia who is currently a DPhil student in the lab of Elizabeth Robertson at the Sir William Dunn School of Pathology, University of Oxford. In fact, you may have already seen her first post as a correspondent in which she covers the Cut + Paste exhibition currently on display in the Francis Crick Institute, London. Alex won the 2020 BSCB Science Writing Prize and here we discuss how she would like to channel her creative energy into the correspondent role. Also, we cover her (international) career path up till now and her love of languages (and word games!).
Congratulations on being selected as one of our new the Node correspondents. How did you hear about the role, and why did you decide to apply for it?
I’ve been following the Node for a while – in fact The Company of Biologists in general – and really like the science- and scientist-centric approach. What I like about the Node is the diversity of the contributors who all have something interesting thing to say. I also enjoy the Tsuku Mogami series of short stories published in Development, which use animated objects to illustrate biological concepts and findings. It’s so creative, and yet so scientific – really inspiring!
So when I saw that the Node was looking for correspondents, I honestly filled in the application within ten minutes or so. I didn’t want to overthink it – if it was meant to be, it was meant to be. I’m thrilled that this turned out to be the case.
Have you done much science writing before, or will this (mostly) be a new experience for you?
Well, not in a very formal way, or within a particular framework. I have written some pieces for my department website (the Sir William Dunn School of Pathology at the University of Oxford). These were either departmental news stories or an overview of papers that were recently published by people working in the department. One thing that perhaps stands out for me is the piece that I submitted for the writing competition of the British Society for Cell Biology. It was three years ago, at the start of my PhD and also at the start of the COVID-19 lockdown, when I had all this energy and very little to do with it. So I poured all that into writing and got really creative with it. I wrote about the evolution of multicellularity and had a lot of fun doing that. It’d be great to bring back some of that creative energy into the correspondent role.
What sort of topics are you excited to write about for the Node? Do they relate to your own work, or are you looking to branch out into other areas?
I’d say that this is a great opportunity to look at something outside of my direct area of research. The ideas that I’ve come up with so far include non-model organism and recent breakthroughs in the medical sciences. As a developmental biologist by training, this will be a unique chance to reach out to people that I normally wouldn’t get to meet.
Actually, I already got to do somethings outside of my normal routine while preparing my first correspondents’ post. I went to report on the Cut + Paste exhibition at the Francis Crick Institute. I’ve never given much thought to what goes into organising an exhibition and it was fun to get a glimpse behind the curtain. I talked to Dr Güneş Taylor, one of the exhibition’s scientific advisors, and heard from the creative consultancy that took on the design of the exhibition. I was the only person there with a notebook, just jotting stuff down. I felt like a real dork, but it was a really fun experience.
Could you tell us a bit more about your own research? What has your career path been like so far?
I was born in Athens and raised in Crete, which is where I went to school. The science teachers at my school, especially for biology and chemistry, were really engaging, which influenced my later choices. I decided to go to Edinburgh for my undergraduate degree, as I had learned all about Dolly the sheep. Besides my scientific education though, I learned that Scotland is just a one-of-a-kind, special place. It’s such a beautiful country to live in!
As part of my undergraduate, I also studied at UC Berkeley for a year, which was a lot of fun. In Edinburgh, though, there was one Professor – Prof. Jamie Davies – who during one of his lectures convinced me that I wanted to study developmental biology. I knew it within the span of 50 minutes, I was like “this is my life now”. I ended up doing my thesis project in his lab, followed by a DPhil (PhD) in Chromosome and Developmental Biology here in Oxford, in Liz Robertson’s lab.
What has it been like to live and work in so many different places?
It’s been absolutely lovely to get the opportunity to make new homes. Of course, it also means that your heart gets broken each time you have to leave again. But then you do always have this second, and third, and fourth home. It’s a really enriching experience and I already find myself thinking where I will go next, and how that will make me feel.
For now, what are you hoping to gain from the experience of being a the Node correspondent?
I’m hoping to meet people with similar interests and see how they approach their job. I’d like to learn from their experience and take some of this back into my own work – my scientific work, but also my writing. My aim is to develop a writing style that is informative and entertaining for as many people as possible. There is already enough literature out there that’s exclusively comprehensible by scientists.
I’d like to think that I’ll always be doing some writing – whether full-time or as a hobby. Ideally, I’ll be working in research for a while longer and being part of the Node will be a great way to make new connections with people I have a lot in common with and whose experience can guide my future steps.
Finally, what do you enjoy doing in your spare time?
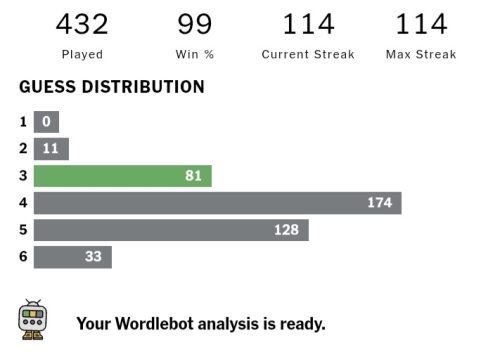
I love word games – I’m currently on a 114-day Wordle streak! I guess it comes from my love of languages. I speak Greek, English, French, Spanish, and ‘baby’ Arabic. I have Arabic-speaking family and I thought it was a good reason for me to learn the language and so – during my first year in Oxford – I would go up to the Center for Islamic Studies for a language class once a week. It was a lot of fun, but it turns out it that I now sound like a cross between a news presenter and a cartoon every time I speak it. It’s because the Arabic they teach you isn’t actually spoken by anyone – it’s the formal version which people don’t use in real life. Still, most people shower you with praise if you can even speak five words in their language, which is always a great feeling.
Our lab is located on the 4th floor of the biomedical building at University of California, Santa Cruz. We are also affiliated with the Institute for the Biology of Stem Cells and Genomics Institute.
Research summary
Ali Shariati: We are interested in understanding unifying principles of stem cell fate decisions during the early days of mammalian development. We also use CRISPR-based approaches to control stem cell fate decisions and assemble them into embryo-like structures.
Lab role call
Gerrald Lodewijk, Postdoctoral Fellow
Research focus: Human embryonic development and implantation, transcriptional mechanisms of G0 to G1 transition in mammalian cell division cycle.
Ben Topacio, Postdoctoral Fellow
Research focus: Determining molecular mechanism linking between cell cycle and cell fate decision in pluripotent stem cells.
Abolfazl Zargaro, PhD student
Research Focus: Development of tools for quantitative analysis of single-cell live microscopy data.
Silvart Arabian, PhD Student
Research Focus: How cells decide to re-enter the cell cycle when they are in G0 phase. Transcriptional mechanisms of G0 to G1 transition in mammalian cell division cycle.
Seungho Lee, PhD Student
Research Focus: How MAPK signalling shape the chromatin landscape and its dynamics in pluripotent stem cells.
Undergraduate researchers: Sayaka Kozuki (Synthetic Models of the embryos), Clara Han (G0 to G1 transition), Lucy Suarez (Cell Cycle Control in pluripotent cells), Arshia Kapil (Microscopy image analysis), Shivani Rao (Microscopy image analysis), Dami Ibrahim (Microscopy Image Analysis).
Favourite technique, and why?
Ali: Biological systems are quite dynamic and time is implicit in biological transitions such as cell fate decisions. So, my favorite method is Single Cell Live Microscopy because it allows us to capture dynamics of cell fate decisions with high temporal resolution. Live microscopy allows us to directly watch individual stem cells as they make important decisions such as to differentiate or not to differentiate. With live microscopy, one has access to information such as morphology of cells (shape, size…), spatial relations of cells and dynamics of signalling pathways that is typically lost in other single cell approaches. Occasionally, you also find cases of individual cells that behave completely different than what is described in the literature.
I am also interested in using CRISPR-based devices to control cell fate and cellular response to signalling cues.
Apart from your own research, what are you most excited about in developmental and stem cell biology
Ali: I am pretty excited about collective cellular decision making that results in reproducible formation of patterns during development as well as application of synthetic biology approaches to control collective behaviour of cells.
How do you approach managing your group and all the different tasks required in your job?
Ali: Managing an academic research laboratory has different aspects such as how to manage projects or how to build a scientifically vibrant environment. One thing I have done is to take an adaptive approach because scientific endeavours are inherently very dynamic in nature and require flexibility in planning. As a result, I am constantly revising my plans and I guess I am also still learning how to lead an academic research laboratory. Periodically, I reflect on how to improve my mentorship and how to promote a dynamic scientific culture that allows for training future generations of scientists with genuine and deep interest in discovering new biology. That last statement is actually one of my professional dreams!
What is the best thing about where you work?
Ali: The campus is so beautiful with many redwood trees, ocean views and wildlife. My colleagues are very supportive and understanding and I also have the chance to work with some very motivated students.
Seungho: Supportive environment, not cutthroat; the opportunity to do research on unanswered questions on stem cells; the Bay area is the centre of innovation in biotech, providing more opportunities to collaborate with other labs or companies.
Silvart: A chance to surround yourself with experts from various research fields. Gain exposure to new technologies and broaden your understanding of stem cells and cell cycle regulation. A supportive environment that fosters growth and development.
Gerrald: Commuting to the lab by bike and getting a free workout! Building our postdoc community (UCSC Postdoc Association).
Ben: I like the lab environment. Everyone is helpful and supportive.
Sayaka: A great opportunity to have research experience during the undergraduate
What’s there to do outside of the lab?
Ali: What I do outside the lab varies a lot. I enjoy (very) long walks with my family, friends or sometimes on my own. I set learning goals for myself such as learning a new language. I also enjoy helping students from resource-limited regions of the Middle East to find their way into higher education.
Seungho: Nature activities; Golf; Santa Cruz doesn’t offer much in terms of nightlife or city activities (great for those who prefer a more peaceful, quiet environment), but San Jose/Palo Alto are not too far away.
Silvart: Sports, playing music and reading books. Spending time outside in nature.
Gerrald: Go to the climbing gym and sauna afterwards. Take a walk or bike ride in one of the nearby state parks.
Ben: Go home. The SF bay area is a short drive away.
Sayaka: Visiting Monterey Aquarium, going Asian supermarkets.
Browse through other ‘Lab meeting’ posts featuring developmental and stem cell biology labs around the world.
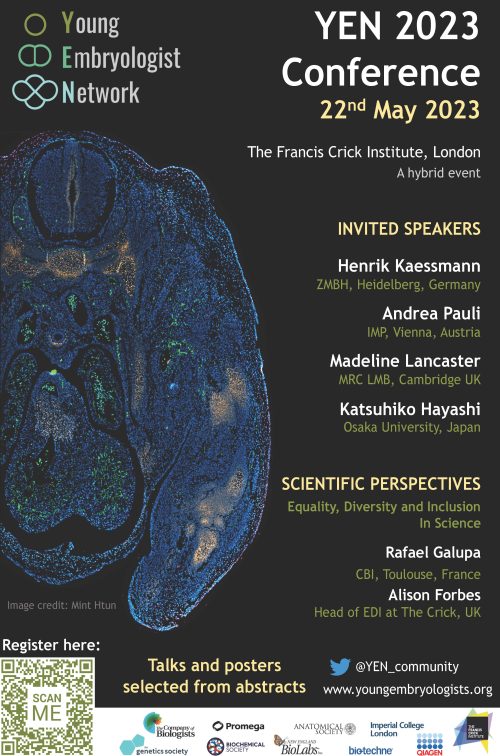
A new edition of the annual Young Embryologist Network conference (YEN 2023) will be hosted at the Francis Crick Institute, in London, on the 22nd of May 2023.
YEN conferences, organised by PhD students and postdocs, have massively increased their impact since virtual attendance was offered in 2021. We are committed to providing an opportunity for early-career developmental biologists to share their research, network, and engage with colleagues in the field. Thus, a hybrid format will also be held this year and attendance will be free thanks to the support of our sponsors.
YEN 2023 will cover some of the most exciting topics in the field, including cross-species comparisons, germ-cell development, and stem-cell-based models of development among others. We have the pleasure of welcoming Andrea Pauli (IMP, Vienna), Madeline Lancaster (MRC LMB, Cambridge), Katsuhiko Hayashi (Osaka University), and Henrik Kaessmann (ZMBH, Heidelberg) as invited speakers. Prof. Henrik Kaessmann will give the Sammy Lee memorial keynote address.
Moreover, the highly acclaimed “Scientific Perspectives” talks are back. This year, the session will focus on “Equality, Diversity and Inclusion in Science”. Rafael Galupa (CBI, Toulouse) and Alison Forbes (Head of the Equality, Diversity & Inclusion committee at the Crick, London) will share their perspectives and experiences on this important subject.
PhD students and postdocs who want to give a short talk or present a poster may submit an abstract by the 20th of April 2023. Registration will be open until the 9th of May 2023.
You can register for this conference, and read a little more about our community, in the link below:
This year’s BSDB Annual General Meeting (AGM) will be held during the Sheffield Spring focus meeting on Cellular Plasticity in Morphogenesis. It will take place on Tuesday the 18th of April, as 16:15 in the main lecture theatre of the conference venue (The Edge). If you would like to attend virtually, please use this link: BSDB AGM Zoom Meeting
Meeting ID: 979 7241 4300
Passcode: dWx@Y7
We would also like to remind you that we will be electing three new BSDB committee members. Please send nominations together with the names of the proposer and seconder to the BSDB Secretary (secretary@bsdb.org) by the 14th of April 2023. Instructions on how to vote will be provided during the AGM.
We look forward to seeing you all there!
Finally, registration opens very soon for the BSDB-hosted European Developmental Biology Congress 25th-28th September 2023. Keep an eye on our website for more info.
If you look at, especially say, identical twins, if one person does testing, you are automatically finding out for your twin what their genetic status is. And so that’s tough, right? Because you’re deciding for them.
Kira Dineen, host of DNA Today
In the latest episode of the Genetics Unzipped podcast, we’re discussing Chris Hemsworth’s recent Alzheimer’s risk findings and the pros and cons of direct-to-consumer DNA testing with genetic counsellor and host of the podcast DNA Today, Kira Dineen.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
 (No Ratings Yet)
(No Ratings Yet) (8 votes)
(8 votes)