As any early career researcher will know, attending your first big scientific meeting is a memorable event in your academic life. Last week, I was lucky enough to attend the Gordon Research Conference in Developmental Biology, held in sleepy South Hadley, two hours outside Boston, Massachusetts. For anyone who wasn’t local, the two hour drive from the airport to the conference site was an ordeal, but the welcome was worth it. The GRC ran an intensive scientific programme of seminars, discussion and poster sessions, held over five days. A Gordon Research Seminar (GRS) also ran over the weekend preceding the conference, exclusively for graduate students and postdocs to interact and present their research in a non-intimidating environment. Sessions spanned kingdoms, with researchers studying both animal and plant models. The themes were varied, and included evo-devo, organoids, and stem cells, as well as the usual suspects such as signalling and gene regulation.
Accommodation building at Mt Holyoke College, South Hadley, MA. This was the venue for the conference this year
In my opinion, running a conference with such a broad title was inspired. For a conference series where meetings often have very specific themes (browse through the list of past events on the GRC website and you will understand what I mean), running an event simply called ‘Developmental Biology’ drew people from all corners of the field. By keeping the title broad, no single mechanism or way of thinking dominated, and in fact several commonalities emerging in the talks were identified in the discussion. For example, the role of mechanical force in development was a recurring mechanism whether you worked on plants, Planarians or people! Perhaps the fact that this theme emerged despite no pre-meditated effort to select talks based around it, means it better reflects where the field is going and what questions are of interest to scientists.
Like at other GRCs, presentation of unpublished work was the star of the show. Speakers of all seniorities did a wonderful job of presenting new and exciting data and hypotheses, making a young researcher like me feel that I was learning things at the cutting edge.
The meeting was also useful to get feedback on published work too.
Publishing a paper can be a lonely experience. You spend years of your life perfecting and revising, then as soon as a manuscript is accepted and released you are left with an eerie silence, a vacuum, while the field digests the data. Apart from a smattering of mentions on social media, post-publication feedback from peers is virtually non-existent for biologists. Yet, when I arrived at the conference it was clear that people had read my paper. Some had even studied for their lab’s journal club! It meant something. Meetings such as the GRC give the community somewhere to exchange ideas. Not only are you there to present your research and absorb the work of others, but also to receive general and specific feedback from scientists across the community- who after all, are your target audience. Many of my most fruitful discussions and interactions happened over breakfast rather than during the seminar sessions themselves.
Several unsurprising topics kept cropping up in talk after talk. Developmental biologists have always been obsessed with pattern formation, and this is certainly not about to change. The field is moving towards computation, and molecular studies are becoming more and more sensitive as everyone who’s anyone seems to be doing single-cell RNA-seq!
The college was surrounded by beautiful wooded countryside and ponds with waterfowl. A peaceful spot to mull over tough scientific questions!
Overall, the atmosphere at the meeting was friendly, fostering open discussion rather than feeling cliquey or competitive. Senior Professors and PhD students were free to interact, and did so. Talking about science, whether it was answering specific questions about my project, or talking about big challenges and how to approach them freed from the shackles of using a particular model organism, technique, or experimental design was fun!
By the end of the week, it is fair to say that the intense schedule began to take its toll, but everyone I spoke to was leaving having learned something, and having been surprised by something. To choose a single stand-out speaker would be impossible, but the quality of the talks were excellent. Even though the meeting was small, with perhaps fewer than one hundred attendees, the GRC did a great job at making researchers across disciplines feel part of the same community. The spirit of the conference was to share ideas, and I came away feeling inspired by people working in areas far removed from my own. After all, as developmental biologists, we are interested in the same fundamental questions, we just use different systems to answer them.
The Schröterlab at the Max Planck Institute of Molecular Physiology in Dortmund is inviting applications for a guest junior scientist (postdoc level).
Our group works in the area of quantitative stem cell biology, asking how cell signaling regulates cell fate decisions at the single cell and the population level. We use embryonic stem cells as an accessible model system, and apply long-term time-lapse imaging in combination with quantitative data analysis to develop predictive models for cell fate decisions. The goal of this project will be to visualize and quantitatively measure cell communication via the FGF/ERK pathway in individual cells, using an embryonic stem cell model for an early fate decision of mammalian embryogenesis (Schröter et al., Development, 2015).
We are looking for recent graduates with a PhD in cell or developmental biology or a related discipline. A quantitative mindset and the willingness to collaborate with theoreticians are essential.
At the Max-Planck-Institute of Molecular Physiology, an international team of scientists from more than 30 nations investigates the basic physical and biochemical processes in cells at the molecular level. The institute is located in the wonderful Ruhr area, the “Ruhrgebiet”. Once famous for its industrial culture and coal mining, in recent years the Ruhrgebiet has transformed into a vibrating and productive science environment. The Ruhr area is home of over 5 million people from over 200 countries, being the largest urban centre of Germany, and the third largest in Europe, after Paris and London. Apart from many theatres and other cultural sites, the region is home to many successful soccer teams, most notably the Dortmund team Borussia.
Those of us who go to the gym are accustomed to thinking of mechanical forces shaping our bodies. Physiological (e.g. determination of bone mass and architecture), pathological (e.g. aneurysm rupture) and even socio-cultural (e.g. lip plates of the Mursi tribe) examples come to mind. The form of most of our organs is templated during embryonic morphogenesis, during which the embryo folds and contorts itself into complex shapes. Origami-like folding of the neuroepithelium to form the neural tube, the embryonic precursor of the central nervous system, has long served as a paradigm of morphogenesis. This process of primary neurulation is fundamentally biomechanical; in mammals it requires progressive tissue-level shape changes to proceed. Failure of these processes, such as insufficient folding of the neuroepithelium at “hinge points”1, are associated with failure of neural tube closure resulting in neural tube defects (NTDs) such as spina bifida. Determining the biomechanics of neural tube closure is therefore a key aim both to understand this fundamental morphogenetic process, and identify causes of NTDs. Here we describe how we have begun to tease apart the biomechanics of neural tube closure in the mammalian spinal region, as recently reported2.
Figure 1 Schematic illustration of a mammalian PNP. The schematic on top is oriented in the tail to head direction as indicated. Schematic cross-sections below illustrate the relationship of the neuroepithelium (NE) to the pre-somitic mesoderm (PM), non-neural ectoderm (NNE) and notochord (NC). The NE bends at a median hinge point (MHP) and paired dorso-lateral hinge points (DLHP). Based on [3] and [2].
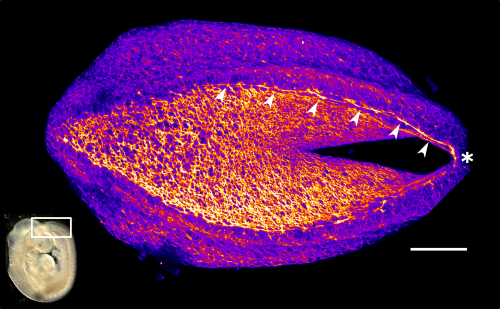
Neural tube closure is a multistep process that requires the coordinated activity of cells of different origins3. It starts with bending of the flat neural plate, creating two opposing neural folds, which progressively elevate and fuse (Figure 1). The fusion process requires the formation of cellular protrusions by non-neural ectoderm cells at the neural fold tips4. These protrusions reach across the midline and “zipper” down the length of the embryo. Zippering reduces the length of the open region of the neural plate, referred to as the posterior neuropore (PNP, Figure 2), as development progresses. PNP length is therefore commonly measured as a readout of the progression of neurulation, and differences in PNP length are extremely useful in identifying developmental stages at which neurulation becomes disrupted. However, in our recent paper we focused on the “perpendicular” process of neural fold apposition, which narrows the PNP. Progressive PNP narrowing also occurs as development progresses, but it has been less intensively studied in recent years.
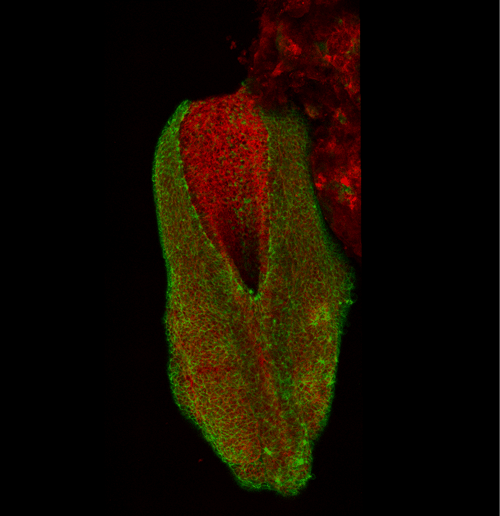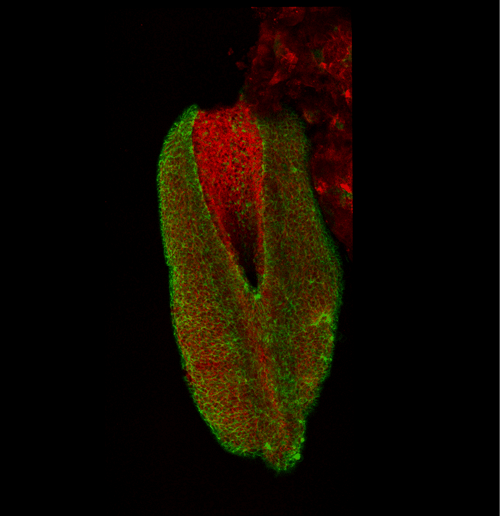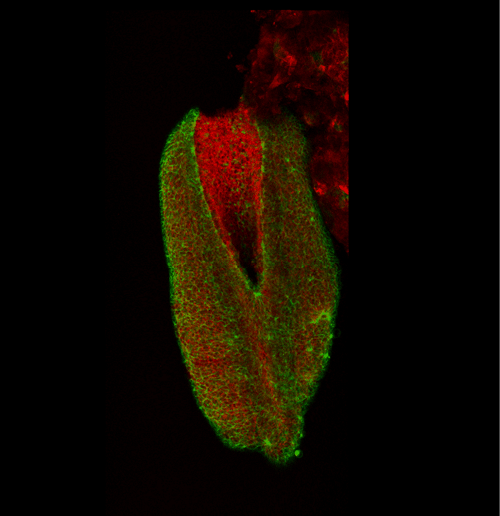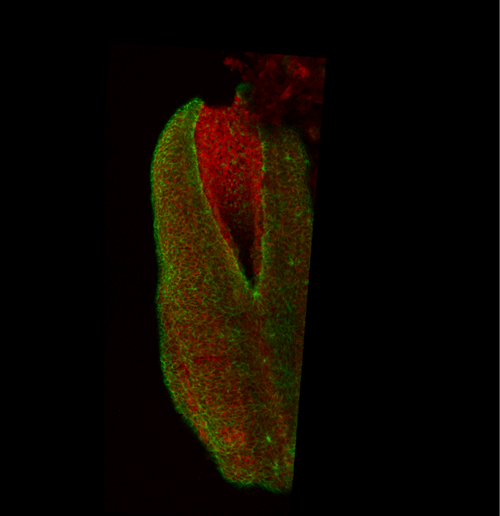
Figure 2 Representative phalloidin-stained mouse PNP showing the F-actin cable (arrow heads) which biomechanically couples the zippering point (*) to the constricting open region. Fire LUT used to highlight F-actin enrichment at the cable and apical surface of the PNP cells. Scale bar = 100 µm for the PNP, the white box on the embryo image inset shows the PNP region.
Whereas significant progress has been made in characterizing the molecular and cellular mechanisms underlying primary neurulation, how these mechanisms biomechanically orchestrate tissue-level shape change is still poorly understood. In our recent review on the topic3 we emphasized that most of the previous work on neurulation biomechanics has been carried out in lower vertebrates and less is known about mammalian neurulation. Lessons learned in simpler models, such as Xenopus5,6 and Ciona7, have substantially advanced our understanding of neurulation as an evolutionarily conserved process. However, extrapolating mechanisms from these models to higher vertebrates must be done with caution given marked differences in the structure of their neural plate and timing over which neurulation occurs3. Thus, our recent paper builds on findings from these simpler models and provides a uniquely mammalian view of the mechanics of primary neurulation.
How the study developed and evolved
Our study started several years ago when Professor Andrew Copp used a glass needle to incise the zippering point in a mouse embryo so as to generate a mechanical model of spina bifida. Unexpectedly, he observed that zippering point incisions caused the neural folds to instantly flip apart, suggesting tension within the surrounding tissue had been pulling them into a more lateral position. This intriguing observation in itself suggested that neural tube closure does not progress through the medial convergence of lateral tissues pushing the folds together as, if that had been the case, incising the zippering point would not have resulted in lateral displacement of the neural folds. Dr Young June Cho, the paper’s second author, bravely took on this finding during his PhD, confirming and expanding the initial observation. However, this model was limited by having to physically incise the neural tube, which carries the potential of inadvertently moving the neural folds. Although physical approaches to testing biomechanics, such as measuring expansion of microsurgical slits8, have been used extensively, we wanted to move towards using less physically invasive methods, namely laser ablation. This became possible in part thanks to the acquisition of a new multiphoton microscope with a Mai Tai laser so powerful it can (accidentally!) set fire to paper, and in part thanks to optimisation of live-embryo positioning and manipulation methods. Laser ablations allowed us to ablate long (>300 µm) lines of tissue from the zippering point along the embryo’s dorsal midline.
This laser ablation method used throughout the paper was set up by the study’s first author, Dr Gabriel Galea, who joined the group on a Wellcome Trust postdoctoral fellowship specifically designed to work on this project. Gabriel had previously collaborated with Prof Copp’s group during his PhD, which was on skeletal cellular mechanobiology. Although the change of fields from bone to neural tube biology was not easy, being a veterinarian he has ample practice in applying knowledge from one situation to another (cats are not small dogs and ferrets are not small cats, but the considerations for treating flea infestations are similar in all three!). The importance of biomechanics to the skeleton is well established: mechanical loading is accepted to be the primary functional determinant of bone mass and architecture. This dogma was established in part thanks to pioneering studies by Gabriel’s PhD supervisors (also vets!), Professors Jo Price and Lance Lanyon. Lance had, many years ago, used strain gauge chips intended for engineering purposes to measure mechanical strains (defined as the percentage change in dimension) in bone. Unfortunately, even the smallest strain gauges are stiffer than neurulation stage embryos, so a non-invasive method was needed to map strains associated with zippering point ablation. Again, engineering provided a solution, in the form of digital image correlation analysis, wherein the relative displacement of pre-placed dots on an object’s surface are used to calculate strains. However implementation of this approach has limitations when applied to biological tissues. Here we turned to a hobby programmer (who also happens to be Gabriel’s father) to script the code for a “Biological Deformation and Strain Measurement” program, quickly renamed “Tissue Deformation and Strain Measurement” (TDSM, which will be available here https://github.com/gauden/tdsm).
After initial validation and testing on simulated datasets, TDSM worked seamlessly when applied to confocal images of mouse PNPs live-imaged before and after zippering point ablation to generate area strain maps. However, the patterns of area strain it showed were so perplexing we described them as “preliminary” even though analyses of sequential embryos all showed the same thing. First of all, they showed long-ranging tissue deformation following laser ablation, far beyond the zippering point, which was unexpected. Secondly, they showed that the tissue around the zippering point itself underwent expansion following ablation. We had confidently assumed the zippering point would be the force-generating structure pulling the neural folds towards the midline, but if that had been the case why would the tissue around it expand following its ablation? Rather, we had expected this region of tissue to retract to a smaller, un-stretched state. Instead we observed tissue constriction far caudally, in the open region of the PNP.
Constriction of the open region suggested a tissue-level mechanism by which the neural folds could be pulled towards the midline. We wanted to see if this tissue constricted during ongoing neurulation but, although we can culture embryos in roller bottle culture for at least 48 hrs, this system precludes visualisation. We therefore reluctantly tried live embryo culture under static conditions while imaging by confocal microscopy, during which the embryos retained a strong heart beat and clear yolk sac circulation. In our setup, even over a relatively short time period (~2 hrs) the embryos underwent substantial morphogenetic movement with very significant narrowing of the posterior neuropore (Video 1, as shown in Galea et al.2). As this happened, the apical surface area of cells in the open region on average decreased, suggesting apical constriction was ongoing. Actomyosin-driven apical constriction is an evolutionarily conserved force-generating mechanism9 and actomyosin is apically enriched in cells of the open neuropore10.
Video 1. Mouse embryo live imaging showing PNP neural fold apposition over ~1 hour, as shown in [2]
This apical actomyosin enrichment had previously been shown in cryostat sections. However, we wanted to visualise it in all its full three-dimensional glory to observe its integration and extent to the zippering point. In so doing we observed that F-actin formed a discrete, continuous cable reaching all the way from the constricting open zone, along the neural folds, up to the zippering point (Figure 2). Laser ablation of the tissue through which this cable runs also results in the neural folds flipping apart, suggesting that the cable biomechanically couples the neuropore. Taking all these findings together produces our working model that the main forces driving neural tube closure are not medial convergence of the surrounding tissue, and not some sort of “pulling” force at the zippering point, but rather constriction of the open neuropore region (Figure 3). The forces generated here are then transmitted to the zippering point by the coupling F-actin cable.
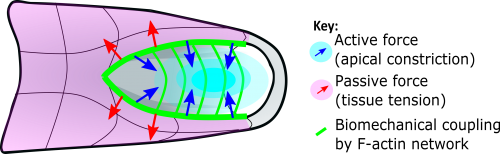
Figure 3 Schematic summary of our findings. Constriction of the NMP zone (cyan) actively generates mechanical force (blue arrows) acting over the length of the PNP biomechanically coupled by the F-actin network (green lines). This opposes stresses within the surrounding tissues (red arrows), facilitating midline apposition of the neural folds, narrowing the zippering point angle and allowing zippering to progress.
Submission and review process
This demonstration of long-ranging biomechanical coupling in a mammalian embryo by an F-actin cable is the main ‘take home message’ of our story, although we do also present further findings in the paper. Given the clinical importance of neural tube closure, we initially submitted our paper to a clinically-focused high impact journal, but were told that although the “editors recognized that [our] developmental biology studies were very well done… these findings would be better suited for a developmental biology-focused venue.” Undeterred, and convinced our findings had broad appeal to a wide readership, we submitted our manuscript to PNAS. Here it received positive reviews which largely requested rewriting sections to make them more accessible. It was actually quite a pleasant experience to feel that addressing reviewers’ comments was improving our manuscript; amending one or two conclusions felt not to be sufficiently robust while generally improving clarity and focus. We particularly appreciated the recognition that our field is still at “the dawn of biomechanics in the early mouse embryo.”
Future outlook
Our ultimate aim in delineating the biomechanics of morphogenesis is to identify and prevent causes of their failure which lead to congenital defects, or to stage-specifically bolster mechanisms which may be deficient in pathological states. In the case of the neural tube, the 3D biomechanical analyses we undertook have fundamentally changed how we think of the progression and completion of spinal neurulation, raising new hypotheses for why they may fail leading to spina bifida. Our own analyses are now expanding beyond the specific roles of the zippering point and adjacent neural folds, to tissues to which they are mechanically coupled several hundred microns away. This new focus is helping us make new cell level observations relevant to the development of spina bifida as well as providing new insights into the integration of signalling cascades and environmental/teratogenic stimuli by biomechanical requirements. Ultimately, as with any research, each question we have answered has raised a dozen more, so watch this space!
Gabriel L Galea and Evanthia Nikolopoulou
UCL Great Ormond Street Institute of Child Health, London, UK
The Department of Integrative Biology (IB) and the Department of Molecular and Cell Biology (MCB) at the University of California, Berkeley are soliciting applications for a 100% (50 % IB, 50% MCB) time new senior level faculty member actively working at the interface between the fields of paleontology and evolutionary developmental biology; this position is open at the tenured level. Potential start date is January 1, 2018 or July 1, 2018.
Through leadership and expertise in the field of paleontology, and the fields of organismal, evolutionary, molecular, cellular, and developmental biology, Berkeley faculty explore questions in adaptation, speciation, ecology, and the genetic and genomic events that have generated organismal diversity. We seek candidates who work at the interface between paleontology and evolutionary developmental biology to leverage the strengths of these fields to address any of a number of questions in evolutionary biology. Areas include (but are not limited to) the evolution of important transitions such as the generation of body plans during the Cambrian, the transition from water to land, the evolution of flight, and the appearance of developmental novelties.
We envision that the position will create synergies for collaborative initiatives in research, teaching and fundraising. The successful candidate will build bridges across disciplines at UC Berkeley (including collaborations with the Departments of Earth and Planetary Sciences, Plant and Microbial Biology and Environmental Science, Policy and Management). S/he will bridge the museum and molecular focused faculty in IB with the molecular and cellular faculty in MCB. Paleontology and Evolutionary Developmental Biology are fields that have great appeal to the public, and an interdisciplinary research scope will provide ample opportunities to highlight the strength of both Departments to the public through outreach programs that could also form the basis for successful fundraising in the future.
Preferred qualifications include demonstrated excellence in research, extensive field or lab experience, evidence of outstanding scholarship within a relevant discipline, a dedication to excellence in teaching at the undergraduate and graduate level, and a commitment to working in an inclusive and interdisciplinary environment. A Ph.D. and/or M.D. or equivalent degree in biology, geology, or a related field is required at the time of application.
Serious consideration will be given to the candidate’s potential for success in mentoring Ph.D. students and teaching at both the undergraduate and graduate levels. We seek someone who combines significant strength in an interdisciplinary program that crosses the Departmental boundaries, which will enhance not only our research standing, but our ability to educate undergraduates and graduate students at the cutting edge of interdisciplinary science. Professional service, including issues of access to and diversity in higher education and the academic profession will also be considered.
Application Procedure:
Applicants who are currently tenured at an institute of higher education/ are an independent investigator must complete an online application via the following link: https://aprecruit.berkeley.edu/apply/JPF0134. If your candidacy progresses, you will be asked to provide contact information for 3-5 referees. We will only contact your referees if you are a finalist for the position, and we will seek your permission before doing so. All letters will be treated as confidential per University of California policy and California state law. Please refer potential referees, including when letters are provided via a third party (i.e., dossier service or career center), to the UC Berkeley statement of confidentiality (http://apo.berkeley.edu/evalltr.html) prior to submitting their letters
All applications should include:
Curriculum Vitae – Your most recently updatedV.
A brief summary of current and future research objectives, teaching interests, and a statement addressing past and/or potential contributions to diversity through research, teaching, and/or service.
The final deadline for applications is July 19, 2017. To receive full consideration, please submit a completed application by this date. Please direct questions to ib_ap_assist@berkeley.edu.
IB and MCB are committed to addressing the family needs of faculty, including dual career couples and single parents. For information about potential relocation to Berkeley, or career needs of accompanying partners and spouses, please visit: http://ofew.berkeley.edu/new-faculty.
The departments seek candidates whose research, teaching, or service has prepared them to contribute to our commitment to diversity and inclusion in higher education. The University of California is an Equal Opportunity/Affirmative Action Employer. All qualified applicants will receive consideration for employment without regard to race, color, religion, sex, sexual orientation, gender identity, national origin, disability, age or protected veteran status. For the complete University of California nondiscrimination and affirmative action policy see: http://policy.ucop.edu/doc/4000376/NondiscrimAffirmAct.
This interview by Aidan Maartens originally appeared in Development, Volume 143, Issue 13.
William ‘Bill’ Harris is Head of the Department of Physiology, Development and Neuroscience at the University of Cambridge, UK, and a Fellow of both the Royal Society and Academy of Medical Sciences. His lab works on the development of the vertebrate nervous system, with a particular focus on cell lineage in the retina. In 2017 he was awarded the British Society for Developmental Biology’s Waddington Medal for outstanding research performance and services to the community. We met Bill in his Cambridge lab to talk science, art and ice hockey.
This year you were awarded the Waddington Medal: what does the award mean to you?
Well, it was a signal that developmental biologists appreciated what I’ve done over my career. I’ve always considered myself first as a neuroscientist, but a developmental neuroscientist, and as I’ve progressed in my career I’ve come to appreciate that the problems I’ve been working on in neuroscience are problems that are common to lots of developmental systems. So to get the appreciation of the field as a whole is very gratifying. It’s the same kind of feeling you get whenever something you’ve done gets accepted, whether a paper or a grant or whatever, that what you’ve done has got through certain hurdles and been deemed OK. Also, as it was near the end of my career and was basically a lifetime achievement award, I was particularly pleased about that.
Let’s go back to the beginning: what got you interested in science, and neuroscience in particular, in the first place?
I got interested in science when I was an undergraduate at the University of California Berkeley, though I guess you could say that before that I was as interested in science as any other kid might be. I did a senior project using one of the early scanning electron microscopes, and decided to look at Drosophila, knowing that no one had ever seen fruit flies blown up to this magnification before. All of a sudden I could see these hidden things, and I knew that geneticists would be interested in them – it was a bit of real, useful science, but was also just amazing to see that level of detail in such a small organism. I keep an image of a Drosophila head from that project up on my office wall to this day.
So that project got me interested in science, but getting interested in the development of the nervous system was a completely different ball game – I got there by a more osmotic, slow process. As a PhD student with Seymour Benzer at CalTech, I was interested in behaviour and genes, in the function of the nervous system. I worked on learning mechanisms in flies, and spent a lot of time in a little dark room getting flies to run to different coloured lights and punishing them if they went to some lights and not others. Then I got some advice that I would spend my whole graduate career just learning about the psychology of flies and not learning any biology if I kept on with that project, so I switched to looking at the fly visual system, and found some mutants that didn’t see certain colours. That got me into the cells, the molecules, the photochemistry and physiology – it was a door into ‘wet’ biology and into the anatomy of the nervous system. That was a better fit for me – I felt I was really learning something.
Seymour Benzer (source), and David Hubel (l) and Torsten Weisel (r) (source)
How did you come to work on the vertebrate nervous system?
As a postdoc I went to Harvard Medical School to work in the labs of David Hubel and Torsten Weisel, who were doing beautiful work on the way the visual cortex operates and the relationship between the physiology of the cortex and its cytoarchitecture. I got interested in the developmental problem of how the early function of the nervous system shapes the way it continues to develop. It was a very exciting time, and the postgrads and postdocs were doing all kinds of interesting, fun experiments. But then one day we were called into the tea room and got a lecture from Hubel and Weisel, who said we shouldn’t just do all these different projects – some of which we had started without even consulting them – and that we were just using their labs as a launching point for our things, whereas they had their own vision for where they wanted the lab to go. They said ‘We’ve planted this plum tree, and now all the plums are very ripe and you guys are just picking them off without even asking!’. Although some of the people were absolutely committed to the visual cortex and didn’t know why they were being told off for doing interesting experiments, it changed my approach to my work – I understood somehow that I should be more independent and that I should try to find my own niche.
So that’s when I switched to a more developmental programme of interests. I moved to axolotls first, largely due to Of Scientists and Salamanders, a book by Victor Twitty, which showed me the wonders of salamander developmental biology. Twitty had found a species of Californian newt that was full of tetrodotoxin, and if you parabiotically joined an embryo of that species to the embryo of an axolotl, the toxin would percolate from the newt embryo (which was insensitive to the toxin) to the axolotl embryo, blocking all of the neural activity in the latter. That allowed me to get back to the earlier question of the role of neural activity in development, which led me to do an experiment revealing that a lot of neural development, including accurate wiring up of the nervous system, did not require any electrical activity. This was an important step forward for the field, which previously thought that neural activity was key to making appropriate connections in the brain. I then started to work on axon guidance – since axons can get to their targets in the absence of neural activity, how exactly do they get there?
So I started with behaviour and have kept going backwards in time until where I am now, which is to ask how different cells in the nervous system get born. That’s probably as far back as I’m going to go or else I’ll move out of the nervous system altogether!
I started with behaviour and have kept going backwards in time until where I am now, which is to ask how different cells in the nervous system get born
Since the early eighties, your lab has mainly focussed on the development of the vertebrate retina. With your wife Christine Holt, you described techniques for labelling neurons to determine lineage and fate – what were the main lessons this early labelling taught you?
In 1980 I started as an Assistant Professor at the University of California San Diego, and a bit later on Christine came to the lab as a postdoc, bringing Xenopuswith her. I used to have a nice life before Christine, in that for the axolotl work you’d get the embryos in the autumn, do your embryonic manipulations, wait for them to grow until you could see what had happened later in the year, then work on writing up these results until the next autumn when the cycle started again. With Xenopus, Christine found a way that you could label specific populations of axons even when they were growing, and not only that, but you could get embryos all year round in the lab – now you could do experiments any day you wanted to and get the results as early as the next day! It was a much more powerful system and allowed the work to go at a faster pace, making the work more exciting and more demanding.
We were interested in the very earliest decisions that retinal ganglion cells made as they were navigating the pathway into the brain. We thought about injecting the cells to label them at earlier and earlier stages, just after they were born, so you could see the first thing the cell did after it was born and knew it was going to be a retinal ganglion cell, to ask how its axon started the journey. We then realised we could inject the cell even before it was born, to look at the mother cell. At the same time, Connie Cepko’s lab had started to use replication-incompetent retroviruses to look at similar problems, and Scott Fraser’s lab had similar ideas, also in Xenopus. We all ended up publishing at around the same time, and had more or less the same results.
The main lesson this labelling taught us was that progenitors in the retina and in the brain were multipotent – they would give rise to many different cell types, ruling out the possibility that there were committed progenitors for each cell type. One thing that puzzled all of us, and led us in the wrong direction for quite a while, was the variability of the lineages – you’d never get the same lineage twice. We all reported this, but didn’t understand it – it might have had to do with different cells experiencing different microenvironments, but it turns out it is a much more intrinsic thing.
In recent years you have had a productive collaboration with Benjamin Simons from the Cavendish Laboratory – how important has mathematical modelling been to your understanding of cell fate in the retina?
It’s shaped the way I think about the variable nature of cell lineages in the retina. Ben came to me after working with Phil Jones on the epidermis of the mouse tail: he was looking at tissues that are homeostatic, where you are losing cells and replacing them at the same rate. Being a physicist he could look at the cell fate data and somehow see in these patterns the underlying model. So he came to me and said that the retina could be a great model to understand how lineages evolve, but I thought that this would probably be too difficult because, during development, things are moving and the rules are going to be changing all the way through. Jie Hie (a postdoc in the lab) and I listened very carefully to the kind of data that he would need, which he could analyse powerfully, and figured out how we might be able to provide it. The analysis revealed that there is very likely a stochastic phase in the development of clones of retinal cells which produces the variability – now we have a model whereby a period of stochasticity means that genes may or may not get expressed, and that this then affects fate in a probabilistic way.
The underlying mathematics isn’t really that hard, but to actually run the simulations and statistically compare the results against the real data is something that Ben is more of an expert at than I. Back when we first published our early clonal data and saw this variability, all I was really able to say was that you can’t discriminate our data from just randomising the numbers of cells that went into the different layers; Ben’s analysis gave us a model that suggests something about the biology. So it really helped to understand those things, and is still helping to motivate work that we’re doing.
In your acceptance speech for the Waddington Medal, you compared the models for cell fate that came out of your work with Waddington’s famous epigenetic landscape. How do Waddington’s ideas relate to your current understanding of stochasticity and determinism in retinal development?
Every developmental biology student has seen Waddington’s epigenetic landscape drawing – it’s a wonderful way to think about development in a diagrammatic way.
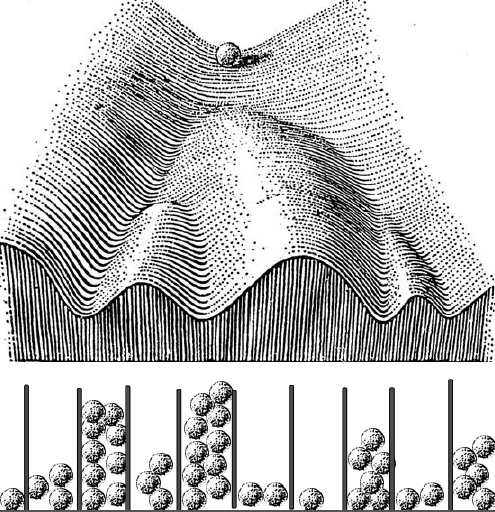
Waddington’s landscape fused with a Galton board, courtesy of Bill Harris
As I was preparing the lecture I wondered what Waddington thought about how cells made decisions to go down one valley or another, but I could not really find much – he described the geography of cell fate decisions, but not how cells made the decisions. His one vague suggestion was that something might be pushing the marbles toward one valley or another. But I was thinking about these probabilities that we’ve been working on, and asked myself if there was a way to join his landscape with probability distributions. As I was thinking about this, my mind went to these Galton boards – you drop a marble, it hits a pin and goes to the right or the left, and then hits another pin and does the same thing, and at the end of that, if you put enough marbles in, you get a beautiful normal distribution in the wells at the bottom. It’s a great way of creating a distribution that is always the same shape even though any particular marble could end up in any of the wells – and if you want to make an organism that’s the same, you want to make a distribution of cell types that’s the same. So let’s run these marbles down through the Waddington landscape, which is really a kind of a Galton board – at each junction it has two choices – but the probabilities are set because of the deepness of the valley and how the ball was rolling. The important thing is that you won’t get the same balls in the same valleys in every organism, but you’ll get the same distribution. So in the talk I just took Waddington’s epigenetic landscape and turned it into a Galton board.
The Waddington Medal lecture can be viewed here
You have been Head of the Department of Physiology, Development and Neuroscience in Cambridge since its inception in 2006. What has it been like steering such a diverse department of researchers?
I came to Cambridge in 1997 as Professor of Anatomy, and the next year I started to take over the reins as Head of Department. In 2006, Anatomy merged with Physiology; there were a lot of similarities in terms of the students we taught and the kinds of research that went on in both departments, so it was a natural union. I think the merger gave a new lease of life to the two departments, and it also helped us with scales of efficiency as we were both running in the red at the time. I don’t consider it a particular challenge to be Head of Department – it’s quite easy if people are motivated to help make the department better. It’s important that everyone has a say in how the place is run – so in meetings I prefer to present ideas and see what everyone thinks, and then together we’ll agree on a way forward democratically.
How did the Cambridge Advanced Imaging Centre (CAIC) come about?
When I came here there was already an installation downstairs that had two electron microscopes, and at the time I was becoming increasingly interested in issues of lineage and taken with making movies of development. We initially applied for a two-photon microscope as a tool to visualise these things. There were clearly advances in light microscopy that were going on and it was important to me that there was a facility in Cambridge that offered top-end microscopy for biologists. I also knew that there were people in the physical sciences at Cambridge that were really interested in advancing microscopy, without necessarily being interested in biological questions. CAIC was an idea that came out of these interactions – we wanted to build a new generation of advanced light microscope instruments specifically for biologists, and quickly tuneable to their specific needs. So in CAIC we build microscopes not just to see how far microscopy can go, but to tackle crucial issues in biology. It has become an amazing facility that a lot of people are using, and is a hub of a wider set of microscopy centres around the university.
You enjoy painting, and in studying the visual system your lab generates beautiful images of retinal organisation. Have you always been a visually oriented person, and has this directed your research?
Since I was a kid I’ve always liked to draw, though I wouldn’t consider myself a particularly talented artist. The science that I’ve done has always depended on imaging – from that very first time when I looked through the scanning electron microscope as an undergraduate, I was hooked on seeing the microscopic world. I can appreciate bands on gels and things like that, but I like to see the cells, and the questions that I’ve ended up asking over the years – like how do axons get from A to B, or how lineages arise – mean that I want to see the processes occurring. I like to see things in three or four dimensions, and the prettier they are the better it is somehow. I particularly love it when a student or postdoc runs into the office saying ‘Bill, you’ve gotta see this!’, and takes me over to the microscope to show me what’s going on.
In my painting – which I turned to again about ten years ago as a serious hobby, in preparation for my retirement – I took some of my doodles, bits of retina and things like that, and started to colour them in with paints. I think I will continue to do biology-themed paintings for a while now.
From that very first time when I looked through the scanning electron microscope as an undergraduate, I was hooked on seeing the microscopic world
I hear you’re into ice hockey? It must be quite a niche pursuit in Cambridge?
In fact, there are quite a few people playing ice hockey in the UK: it’s a niche sport but it’s growing. I grew up as a Canadian, and was on skates when I was four or five – we used to flood our back yard to make an ice rink, and I would play ice hockey with my brothers. I was never a great player, like I’m not a great painter, but I liked it enough that I kept doing it. When I came here to Cambridge, I discovered there was a team – in fact, the oldest ice hockey rivalry in the world is between Oxford and Cambridge. So I quickly got involved, became the coach of the team, and also play in the recreational team, though there I’m more of a hazard than a help.
Is there anything that Development readers would be surprised to find out about you?
I am the youngest of four children – an older sister and two older brothers. So I was the baby of the family and have always been treated that way. I keep thinking that after all my years of experience as Head of Department, and in life more generally, I’m going to meet them one day and be able to hold my own. But no, they still treat me as a useless baby brother who doesn’t know anything and is not worth listening to. After all these years, it hasn’t changed!
Our latest monthly trawl for developmental biology (and other cool) preprints. See last year’s introductory post for background, and let us know if we missed anything
We started this feature a year ago, and each month it’s taken a little longer to curate as more labs decide to deposit preprints alongside conventional channels of publication, and as this practise is encouraged by an increasing number of journals, funding bodies and academic organisations.
Highlights this anniversary month include a lot of neural development, analyses of a controversial CRISPR off target paper (and a word from the authors of the original paper), an analysis of stem cells and mutation in long lived trees, a slew of new genomes (gar, greyling, reindeer, zucchini and colonial tunicate!), and the routine acronym-fest of new data analysis tools (incorporating mimosas and pineapples this time). The preprints were hosted on bioRxiv, PeerJ and arXiv.
Genome Architecture Leads a Bifurcation in Cell Identity. Sijia Liu, Haiming Chen, Scott Ronquist, Laura Seaman, Nicholas Ceglia, Walter Meixner, Lindsey A. Muir, Pin-Yu Chen, Gerald Higgins, Pierre Baldi, Steve Smale, Alfred Hero, Indika Rajapakse
Epigenetic resetting of human pluripotency. Ge Guo, Ferdinand von Meyenn, Maria Rostovskaya, James Clarke, Sabine Dietmann, Duncan Baker, Anna Sahakyan, Samuel Myers, Paul Bertone, Wolf Reik, Kathrin Plath, Austin Smith
Low Rate of Somatic Mutations in a Long-Lived Oak Tree. Namrata Sarkar, Emanuel Schmid-Siegert, Christian Iseli, Sandra Calderon, Caroline Gouhier-Darimont, Jacqueline Chrast, Pietro Cattaneo, Frederic Schutz, Laurent Farinelli, Marco Pagni, Michel Schneider, Jeremie Voumard, Michel Jaboyedoff, Christian Fankhauser, Christian S. Hardtke, Laurent Keller, John R. Pannell, Alexandre Reymond, Marc Robinson-Rechavi, Ioannis Xenarios, Philippe Reymond
Transgenic flatworms from Wudarksi, et al.’s preprint
A platform for efficient transgenesis in Macrostomum lignano, a flatworm model organism for stem cell research. Jakub Wudarski, Daniil Simanov, Kirill Ustyantsev, Katrien de Mulder, Margriet Grelling, Magda Grudniewska, Frank Beltman, Lisa Glazenburg, Turan Demircan, Julia Wunderer, Weihong Qi, Dita B. Vizoso, Philipp M. Weissert, Daniel Olivieri, Stijn Mouton, Victor Guryev, Aziz Aboobaker, Lukas Scharer, Peter Ladurner, Eugene Berezikov
Chd8 haploinsufficient mice display anomalous behaviours, increased brain size and cortical hyper-connectivity. Philipp Suetterlin, Shaun Hurley, Conor Mohan, Kimberley L. H. Riegman, Marco Pagani, Angela Caruso, Jacob Ellegood, Alberto Galbusera, Ivan Crespo-Enriquez, Caterina Michetti, Robert Ellingford, Olivier Brock, Alessio Delogu, Philippa Francis-West, Jason P. Lerch, Maria Luisa Scattoni, Alessandro Gozzi, Cathy Fernandes, Albert Basson
Evolutionary proteomics uncovers ciliary signaling components. Monika Abedin Sigg, Tabea Menchen, Jeffery Johnson, Chanjae Lee, Semil P. Choksi, Galo Garcia III, Henriette Busengdal, Gerard Dougherty, Petra Pennekamp, Claudius Werner, Fabian Rentzsch, Nevan Krogan, John B. Wallingford, Heymut Omran, Jeremy F. Reiter
Cell biology
The β3-integrin endothelial adhesome regulates microtubule dependent cell migration. Samuel J. Atkinson, Aleksander M. Gontarczyk, Tim S. Ellison, Robert T. Johnson, Benjamin M. Kirkup, Abdullah Alghamdi, Wesley J. Fowler, Bernardo C. Silva, Jochen J. Schneider, Katherine N. Weilbaecher, Mette M. Mogensen, Mark D. Bass, Dylan R. Edwards, Stephen D. Robinson
BPG: Seamless, Automated and Interactive Visualization of Scientific Data. Christine P’ng, Jeffrey Green, Lauren C. Chong, Daryl Waggott, Stephenie D. Prokopec, Mehrdad Shamsi, Francis Nguyen, Denise Y. F. Mak, Felix Lam, Marco A. Albuquerque, Ying Wu, Esther H. Jung, Maud H. W. Starmans, Michelle A. Chan-Seng-Yue, Cindy Q. Yao, Bianca Liang, Emilie Lalonde, Syed Haider, Nicole A. Simone, Dorota Sendorek, Kenneth C. Chu, Nathalie C. Moon, Natalie S. Fox, Michal R. Grzadkowski, Nicholas J. Harding, Clement Fung, Amanda R. Murdoch, Kathleen E. Houlahan, Jianxin Wang, David R. Garcia, Richard de Borja, Ren X. Sun, Xihui Lin, Gregory M. Chen, Aileen Lu, Yu-Jia Shiah, Amin Zia, Ryan Kearns, Paul Boutros
Persistent homology demarcates a leaf morphospace. Mao Li, Hong An, Ruthie Angelovici, Clement Bagaza, Albert Batushansky, Lynn Clark, Viktoriya Coneva, Michael Donoghue, Erika Edwards, Diego Fajardo, Hui Fang, Margaret Frank, Timothy Gallaher, Sarah Gebken, Theresa Hill, Shelley Jansky, Baljinder Kaur, Philip Klahs, Laura Klein, Vasu Kuraparthy, Jason Londo, Zoe Migicovsky, Allison Miller, Rebekah Mohn, Sean Myles, Wagner Otoni, J. Chris Pires, Edmond Riffer, Sam Schmerler, Elizabeth Spriggs, Christopher Topp, Allen Van Deynze, Kuang Zhang, Linglong Zhu, Braden M. Zink, Daniel H. Chitwood
Critical Assessment of Metagenome Interpretation − a benchmark of computational metagenomics software. Alexander Sczyrba, Peter Hofmann, Peter Belmann, David Koslicki, Stefan Janssen, Johannes Droege, Ivan Gregor, Stephan Majda, Jessika Fiedler, Eik Dahms, Andreas Bremges, Adrian Fritz, Ruben Garrido-Oter, Tue Sparholt Jorgensen, Nicole Shapiro, Philip D Blood, Alexey Gurevich, Yang Bai, Dmitrij Turaev, Matthew Z DeMaere, Rayan Chikhi, Niranjan Nagarajan, Christopher Quince, Fernando Meyer, Monika Balvociute, Lars Hestbjerg Hansen, Soren J Sorensen, Burton K H Chia, Bertrand Denis, Jeff L Froula, Zhong Wang, Robert Egan, Dongwan Don Kang, Jeffrey J Cook, Charles Deltel, Michael Beckstette, Claire Lemaitre, Pierre Peterlongo, Guillaume Rizk, Dominique Lavenier, Yu-Wei Wu, Steven W Singer, Chirag Jain, Marc Strous, Heiner Klingenberg, Peter Meinicke, Michael Barton, Thomas Lingner, Hsin-Hung Lin, Yu-Chieh Liao, Genivaldo Gueiros Z Silva, Daniel A Cuevas, Robert A Edwards, Surya Saha, Vitor C Piro, Bernhard Y Renard, Mihai Pop, Hans-Peter Klenk, Markus Goeker, Nikos C Kyrpides, Tanja Woyke, Julia A Vorholt, Paul Schulze-Lefert, Edward M Rubin, Aaron E Darling, Thomas Rattei, Alice C McHardy
Accessible, curated metagenomic data through ExperimentHub. Edoardo Pasolli, Lucas Schiffer, Paolo Manghi, Audrey Renson, Valerie Obenchain, Duy Tin Truong, Francesco Beghini, Faizan Malik, Marcel Ramos, Jennifer B Dowd, Curtis Huttenhower, Martin Morgan, Nicola Segata, Levi Waldron
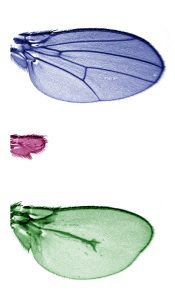
The top and bottom of the image show how the Dpp concentration gradient affects the organisation of the wing structure of Drosophila melanogaster. In the centre, in the absence of Dpp, the wing does not grow. Image: Lara Barrio.
• Scientists at IRB Barcelona clarify the function of the genes that drive wing development in the fruit fly Drosophila melanogaster.
• Published in the journal eLife, this study unveils that the Dpp morphogen is necessary for wing growth but that its gradient does not govern this process.
• Understanding the development of limbs in Drosophila paves the way to research into congenital defects in vertebrates.
Barcelona, 5th July 2017.- Researchers working in the Development and Growth Control Lab at IRB Barcelona reveal that the Dpp gene (BMP in humans) plays a double role in the structural organisation and growth of the wings of the fruit fly Drosophila melanogaster.
This study, which has been published in the journal eLife, demonstrates that Dpp is necessary for tissue growth but that “its gradient does not direct wing growth,” explains Marco Milán, ICREA research professor and head of the study. This and two other studies published simultaneously in the journal eLife settle the intense scientific debate regarding the function of Dpp and other morphogens involved in development.
Morphogens are molecules found in concentration gradients throughout tissues and they send signals from one cell to another. “The wing of Drosophila melanogaster has several morphogens, such as Dpp (BMP in humans) and Wingless (Wnt in humans), which are necessary for growth,” explains Lara Barrio, the first author of the study and postdoctoral fellow in the Development and Growth Control Lab at IRB Barcelona. In this study, the scientists have examined how Dpp regulates growth and analysed how cells behave when Dpp levels are manipulated.
The role of the Dpp concentration gradient in the regulation of tissue is the subject of intense debate among scientists. Morphogens have been considered to be responsible for this process; however, using distinct techniques, these three studies now conclude that morphogens are necessary for growth but that their concentration gradients do not directly govern this process.
“We know that the gradient of this morphogen in particular affects the structural organisation or the identity of the tissue, but the different levels of Dpp across the tissue have no effect on growth. That is to say, whether a tissue grows or not depends on whether Dpp is present or absent. Its gradient has no influence”, explains Marco Milán.
So what regulates the size of the final structural of the Drosophila wing? “Morphogen gradients don’t. There must be another alternative and as yet unknown mechanism, and the fly wing is an ideal model to answer this question,” says Marco Milán.
This research is consistent with knowledge about the morphogen Sonic hedgehog in vertebrate limbs. The gradients of this molecule affect tissue identity (for example, for the fingers of a hand to differ from each other) but do not regulate growth. Therefore understanding how the structures of Drosophila form and develop paves the way to studying vertebrate development and congenital defects in humans.
This study has been supported by the Ministry of Economy, Industry and Competitiveness (MINECO), ERDF “Una manera de hacer Europa”, and the CERCA Programme of the Government of Catalonia.
Reference article:
Lara Barrio and Marco Milán
Boundary Dpp promotes growth of medial and lateral regions of the Drosophila wing
eLife (2017). DOI: 10.7554/eLife.22013
Post-doc in mathematical modeling on phenotypic evolution and embryonic development:
1.Job/ project description:
The postdoc could choose between three main research projects:
a. Mathematical modeling of phenotypic evolution in populations with embryonic development.
b. Mathematical modeling of gene network and embryonic development evolution.
c. Mathematical modeling of organ development and their evolution in mammalian teeth or Drosophila wing.
The actual project will be chosen together with the candidate depending on his/her interests and skills.
The research will take place in the Isaac Salazar-Ciudad’s group in the Center of Excellence in Experimental and computational developmental biology of the Biotechnology Institute of the University of Helsinki, Finland.
The job is for 1 year and renewable for 1 extra year.
2. Background:
Why organisms are the way they are?
Can we understand the processes by which complex organisms are build in each generation and how these evolved?
We are trying…
The process of embryonic development is now widely acknowledged to be crucial to understand evolution since any change in the phenotype in evolution (e.g. morphology) is first a change in the developmental process by which this phenotype is produced. Over the years we have come to learn that there is a set of developmental rules that determine which phenotypic variation can possibly arise in populations due to genetic mutation (the so called genotype-phenotype map). Since natural selection can act only on existing phenotypic variation, these rules of development have an effect on the direction of evolutionary change.
Salazar-Ciudad’s group is devoted to understand these developmental rules and how these can help to better understand the direction of evolutionary change. The ultimate goal is to modify evolutionary theory by considering not only natural selection in populations but also developmental biology in populations. For that aim we combine mathematical models of embryonic development that relate genetic variation to morphological variation with population models. The former models are based on what is currently known in developmental biology.
Salazar-Ciudad’s group is in close collaboration with Jukka Jernvall’s group and other groups within the center of excellence in experimental and computational developmental biology. The center includes groups working in tooth, wing, hair and mammary glands development. In addition to evolutionary and developmental biologists the center of excellence includes bioinformaticians, populational and quantitative geneticists, systems biologists and paleontologists.
“The Academy of Finland’s Centres of Excellence are the flagships of Finnish research. They are close to or at the very cutting edge of science in their fields, carving out new avenues for research, developing creative research environments and training new talented researchers for the Finnish research system.”
3. Requirements:
The applicant must hold a PhD in either evolutionary biology, developmental biology or, preferably, in evolutionary developmental biology (evo-devo). Applicants with a PhD in theoretical or mathematical biology are also welcome.
Programming skills or a willingness to acquire them is required.
The most important requirement is a strong interest and motivation on science and evolution. A capacity for creative and critical thinking is also required.
4. Description of the position:
The fellowship will be for a period of up to 1+1 years (100% research work: no teaching involved).
Salary according to Finnish postdoc salaries.
5. The application must include:
-Motivation letter including a statement of interests
-CV (summarizing degrees obtained, subjects included in degree and grades, average grade).
-Summary of PhD project, its main conclusions and its underlying motivation.
-Application should be sent to Isaac Salazar-Ciudad by email:
isaac.salazar@helsinki.fi
No official documents are required for the application first stage but these may be required latter on.
6. Deadline:
There is no specific deadline, the position will be filled as soon as a suitable candidate is found.
7. Examples of recent publications by Isaac Salazar-Ciudad group.
Brun-Usan M, Marín-Riera M, Grande C, Truchado-Garcia M, Salazar-Ciudad I. A set of simple cell processes is sufficient to model spiral cleavage. Development. 2017 Jan 1;144(1):54-62.
-Salazar-Ciudad I, Marín-Riera M. Adaptive dynamics under development-based genotype-phenotype maps. Nature. 2013 May 16;497(7449):361-4.
-Salazar-Ciudad I, Jernvall J. A computational model of teeth and the developmental origins of morphological variation. Nature. 2010 Mar 25;464(7288):583-6.
8. Interested candidates should check our group webpage:
Here are the highlights from the current issue of Development:
Heart tube formation: a gut reaction
Morphogenesis of the endoderm-derived foregut (FG) is tightly linked to that of the mesoderm-derived heart tube (HT), with both structures arising at approximately the same time and place in the developing embryo. However, the physical forces that create the FG and HT are unclear. Here, Larry Taber and colleagues combine experimental approaches in chick embryos with computational modelling to explore this issue (p. 2381). They propose that differential anisotropic growth between the mesoderm and endoderm drives tissue folding and formation of the FG while also bringing the bilateral heart fields (HFs) into close proximity. Indeed, inhibition of cell proliferation (using the mitotic inhibitor Aphidicolin) together with computational simulations confirm that proliferation is required for this initial step. They further propose that actomyosin contraction in the anterior intestinal portal (AIP; the caudal opening of the FG) then generates tension that elongates the FG and the fused HFs. In line with this, inhibition of contraction (using the myosin inhibitor blebbistatin) and modelling analyses reveal that contraction is required for FG and HF elongation. Finally, the authors reveal that the fused HFs thicken and expand – driven by an accumulation of cardiac jelly – to eventually create the HT. Together, these findings highlight a new model that integrates HT and FG morphogenesis.
mTORC-ing some sense into pancreas development
In recent years, much progress has been made in uncovering the signalling pathways and transcriptional networks that can influence pancreas development during embryogenesis. However, comparatively little is known about postnatal development of the pancreas, and whether nutrients can impact pancreas development and function after birth. Now, James Wells, Katie Sinagoga and colleagues demonstrate that the nutrient-sensing mTOR pathway regulates the maturation and function of mouse pancreatic islets postnatally (p. 2402). They first reveal that mTOR is dispensable for embryonic development but is required for normal postnatal islet development, with levels of mTOR signalling being highest in the first few weeks after birth. The researchers further report that deletion of mTOR in the endocrine pancreas causes a decrease in islet mass and compromised islet maturation and morphogenesis; this is accompanied by a decrease in islet function. Finally, the authors show that the two known mTOR-containing complexes – mTORC1 and mTORC2 – mediate distinct functions of mTOR: while mTORC1 predominantly regulates islet maturation and function, mTORC2 influences islet mass and morphogenesis. Overall, these findings highlight a potential role for nutrient-sensing mechanisms during postnatal islet development and maturation, a finding that has important implications for deriving functional β-cells in vitro.
A role for cell repulsion during placental labyrinth formation
The placental labyrinth – a complex structure made up of trophoblasts and endothelial cells – provides the interface for gas and nutrient exchange between the embryo and the mother and hence is essential for embryogenesis. However, the molecular mechanisms that underlie the development of this vital labyrinth, particularly those that influence its vascularization, are poorly understood. Here, on p. 2392, Yoshiaki Kubota, Satoru Yamagishi and co-workers report the unexpected finding that fibronectin leucine-rich transmembrane protein 2 (FLRT2), which is a protein that acts as a chemorepellent in neurons, regulates placental labyrinth development in mice. They report that FLRT2 is expressed in endothelial cells specifically in the placental labyrinth. The researchers further demonstrate that the vasculature is poorly formed and aberrantly organized in FLRT2-deficient placentas, with FLRT2-deficient embryos exhibiting high levels of hypoxia. In vitro assays reveal that, as occurs in neurons, FLRT2 signals through UNC5B and can mediate cell repulsion. Following on from this, the authors show that Unc5b deletion recapitulates the vascular defects observed in Flrt2-deficient placentas. Together these exciting results point towards a role for inter-endothelial repulsion, mediated by FLRT2, during placental morphogenesis.
PLUS…
An interview with Bill Harris
William ‘Bill’ Harris is Head of the Department of Physiology, Development and Neuroscience at the University of Cambridge, UK, and a Fellow of both the Royal Society and Academy of Medical Sciences. His lab works on the development of the vertebrate nervous system, with a particular focus on cell lineage in the retina. In 2017 he was awarded the British Society for Developmental Biology’s Waddington Medal for outstanding research performance and services to the community. We met Bill in his Cambridge lab to talk science, art and ice hockey. Read the Spotlight on p. 2307
MicroRNAs in neural development: from master regulators to fine-tuners
The proper formation and function of neuronal networks is required for cognition and behavior. Indeed, pathophysiological states that disrupt neuronal networks can lead to neurodevelopmental disorders such as autism, schizophrenia or intellectual disability. In recent years, it has been shown that microRNAs (miRNAs), an abundant class of small regulatory RNAs, can regulate neuronal circuit development, maturation and function by controlling, for example, local mRNA translation. Here, Marek Rajman andGerhard Schratt provide an overview of the most prominent regulatory miRNAs that control neural development, highlighting how they act as ‘master regulators’ or ‘fine-tuners’ of gene expression, depending on context. See the Review on p. 2310
Human haematopoietic stem cell development: from the embryo to the dish
Haematopoietic stem cells (HSCs) emerge during embryogenesis and give rise to the adult haematopoietic system. Understanding how early haematopoietic development occurs is of fundamental importance for basic biology and also for recapitulating the development of HSCs from pluripotent stem cells in vitro. Here, Alexander Medvinsky and colleagues discuss what is known of human haematopoietic development: the anatomical sites at which it occurs, the different temporal waves of haematopoiesis, the emergence of the first HSCs and the signalling landscape of the haematopoietic niche. They also discuss the extent to which in vitro differentiation of human pluripotent stem cells recapitulates bona fide human developmental haematopoiesis, and outline some future directions in the field. See the Review on p. 2323
I am Eleni Chrysostomou, a PhD student in Uri Frank‘s lab at the National University of Ireland, Galway. The Frank lab’s general interest is development and regeneration, stem and germ cell biology, neural fate commitment, and the chromatin biology underlying these processes. The focus of my project is the roles of SoxB transcription factors (TFs) during nervous system development and regeneration. More specifically, my hypothesis is that SoxB TFs are expressed sequentially in the neural lineage and play a role in neural progenitor cells (NPCs) migration from the body column to the site of injury to re-establish and regenerate the missing head region. The work is mostly done in an in vivo context utilizing transgenic animals, as well as various molecular techniques.
According to the Greek mythology, one of Hercules’ labours was to kill the sea monster Lernaean hydra. What he didn’t know was that every time he decapitated one of the monster’s heads, it would grow back in triplicate! But how is that even remotely related to stem cell biology…
With that said, meet our animal model Hydractinia. Hydractinia, a marine colonial hydrozoan can be described as a great representative of the Cnidaria phylum and an excellent animal model to study cell and developmental biology, as its utility let to the assembly of the very first concepts and terms in biology, including the characterization of stem cells (Weismann, 1883).
The stem cells founded in Hydractinia (aka interstitial cells: i-cells) remain collectively pluripotent throughout the organism’s life and they express germ line markers such as Nanos, Vasa and Piwi (Bradshaw et al., 2015; Plickert et al., 2012). Hydractinia can be easily cultured and genetically manipulated in the laboratory without any ethical restrictions, and its application is suitable in various disciplines.
Hydractinia has a relatively simple life cycle. Following fertilization, the embryonic development lasts 36-48 hours and upon induced metamorphosis the primary polyp is asexually reproduced to give rise to the new colony (Figure 1). The resulting members of an individual colony share a gastrovascular space, nervous system and migratory stem cells by maintaining tissue continuity (Gahan et al., 2016)
Figure 1: The life cycle of Hydractinia (Image obtained from Flici et al., 2017, Cell Reports).
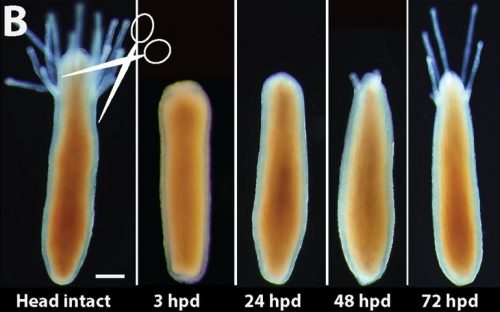
What really makes this animal so intriguing is its regenerative abilities. Like the mythical creature Lernaea, upon decapitation the animal is able to regenerate a fully functional and complete head just within three days (Figure 2).
Figure 2: Live imaging of head regeneration in Hydractinia (Image obtained from Bradshaw et al., 2015, eLife).
What do we do with our “non-mainstream” animal model…
The Frank Lab is based in the National University of Ireland Galway and it is composed of post-graduate students and Post-docs with each of us exploring a different aspect of the biology of these animals in order to answer fundamental questions spanning from developmental biology to epigenetics.
Some of the projects that are currently running in our lab are: whole body regeneration from small tissue fragments (Hakima Flici – Post-Doc), the role of Piwi genes in development and regeneration (Emma McMahon – PhD student), histone variants in Hydractinia (Anna Torok – PhD student), sexual commitment (Timothy Dubuc – Post-Doc).
A typical day in our lab is anything but ordinary. We start the day by spawning the animals and collect the embryos for injection. As the time window for injection is quite narrow – only half an hour before they start dividing – you don’t have much of a choice but to wake up and run downstairs to the manipulation room. After the running and once injection is done, you can breathe for a bit but not for long. The animals need to be fed after all that effort given to the spawning. Generally, there is not much time to lay back and enjoy a nice cup of coffee – not a tea person – but we still love it (that’s why we are in research I guess). Once the feeding is done, you can go back to the lab and see what’s the plan for the day and continue with the experiments that you left from last night – most of the protocols have overnight incubations or you were just too tired and hungry to keep working, lets be honest. And before you realize it it’s already late in the afternoon! The working hours in Ireland can be a bit tricky during the summer time (not really summer, but oh well) as the day light can last until 10-11 o’clock in the night and you have no sense of the actual time! The supervisors are happy during that period…
Even though working with a not so popular animal model can be a bit demanding, it gives us the opportunity to study developmental aspects not feasible in other animal models. Although structurally simple, Hydractinia encompasses a complex gene repertoire that is highly conserved to their sister branch bilaterians. Not many organisms are able to regenerate body parts and especially their heads. This pioneering model for stem cell biology gives as a highly valuable advantage to gain insights on why other animals including humans have limited or zero abilities to regenerate missing parts of their bodies.
The Frank Lab. Some people missing as we never manage to get a picture with all of us.


 (7 votes)
(7 votes) (No Ratings Yet)
(No Ratings Yet)
 (2 votes)
(2 votes)











 William ‘Bill’ Harris is Head of the Department of Physiology, Development and Neuroscience at the University of Cambridge, UK, and a Fellow of both the Royal Society and Academy of Medical Sciences. His lab works on the development of the vertebrate nervous system, with a particular focus on cell lineage in the retina. In 2017 he was awarded the British Society for Developmental Biology’s Waddington Medal for outstanding research performance and services to the community. We met Bill in his Cambridge lab to talk science, art and ice hockey. Read the Spotlight on p.
William ‘Bill’ Harris is Head of the Department of Physiology, Development and Neuroscience at the University of Cambridge, UK, and a Fellow of both the Royal Society and Academy of Medical Sciences. His lab works on the development of the vertebrate nervous system, with a particular focus on cell lineage in the retina. In 2017 he was awarded the British Society for Developmental Biology’s Waddington Medal for outstanding research performance and services to the community. We met Bill in his Cambridge lab to talk science, art and ice hockey. Read the Spotlight on p.  The proper formation and function of neuronal networks is required for cognition and behavior. Indeed, pathophysiological states that disrupt neuronal networks can lead to neurodevelopmental disorders such as autism, schizophrenia or intellectual disability. In recent years, it has been shown that microRNAs (miRNAs), an abundant class of small regulatory RNAs, can regulate neuronal circuit development, maturation and function by controlling, for example, local mRNA translation. Here,
The proper formation and function of neuronal networks is required for cognition and behavior. Indeed, pathophysiological states that disrupt neuronal networks can lead to neurodevelopmental disorders such as autism, schizophrenia or intellectual disability. In recent years, it has been shown that microRNAs (miRNAs), an abundant class of small regulatory RNAs, can regulate neuronal circuit development, maturation and function by controlling, for example, local mRNA translation. Here,  Haematopoietic stem cells (HSCs) emerge during embryogenesis and give rise to the adult haematopoietic system. Understanding how early haematopoietic development occurs is of fundamental importance for basic biology and also for recapitulating the development of HSCs from pluripotent stem cells in vitro. Here, Alexander Medvinsky and colleagues discuss what is known of human haematopoietic development: the anatomical sites at which it occurs, the different temporal waves of haematopoiesis, the emergence of the first HSCs and the signalling landscape of the haematopoietic niche. They also discuss the extent to which in vitro differentiation of human pluripotent stem cells recapitulates bona fide human developmental haematopoiesis, and outline some future directions in the field. See the Review on p.
Haematopoietic stem cells (HSCs) emerge during embryogenesis and give rise to the adult haematopoietic system. Understanding how early haematopoietic development occurs is of fundamental importance for basic biology and also for recapitulating the development of HSCs from pluripotent stem cells in vitro. Here, Alexander Medvinsky and colleagues discuss what is known of human haematopoietic development: the anatomical sites at which it occurs, the different temporal waves of haematopoiesis, the emergence of the first HSCs and the signalling landscape of the haematopoietic niche. They also discuss the extent to which in vitro differentiation of human pluripotent stem cells recapitulates bona fide human developmental haematopoiesis, and outline some future directions in the field. See the Review on p. 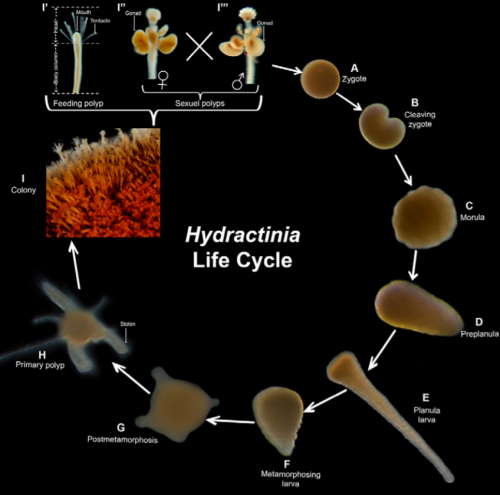

{kind=link}