Applications are still open for the Comparative Invertebrate Embryology course at the Friday Harbor Labs. It’s a great opportunity to work with diverse animal embryos. We attempt to provide an integrated view of animal development, bridging cell and molecular mechanisms with ecological and evolutionary processes. Marine invertebrate development can provide new perspectives on developmental mechanisms, eco-devo, evo-devo, regeneration, and tissue engineering.
Applications are needed before Feb. 29th. Financial aid may be available.
The course runs from June 13 – July 15, 2016 (5 weeks).
— Image: Friday Harbor Labs —
The Friday Harbor Labs give access to an extraordinary variety of organisms from diverse marine habitats, and provide opportunities to interact with a broad community of biologists.
The focus of the course is hands-on observation of living embryos and larvae from over a dozen animal phyla. Techniques students will learn may* include:
Spawning and culture of embryos and larvae from diverse phyla.
Confocal microscopy and SEM
Microinjection
Methods for measuring cell/embryo mechanical properties
Instructors include Dr. Sally Leys and myself. Dr. Leys is an expert on sponge development and the evolution of animal body plans. My research focuses on the roles of biomechanics in development-environment interactions. We will also have guest lectures from experts on diverse topics and taxa, including Drs. Brad Shuster, Richard Strathmann, Sophie George, Tony Pires, and others TBD.
The Media Fellowships provide a unique opportunity for practising scientists, clinicians and engineers to spend two to six weeks working at the heart of a media outlet such as the Guardian, BBC Breakfast or the Londonist.
Every year up to ten Media Fellows are mentored by professional journalists and learn how the media operates and reports on science, how to communicate with the media and to engage the wider public with science through the media.
After their media placement Fellows attend the British Science Festival in September, which provides an opportunity to gain valuable experience working alongside a range of media organisations from all over the UK in our dedicated Press Centre. The Festival also offers opportunities to learn from a wide range of public engagement activities and network with academics, journalists and science communicators.
A 3-year PhD studentship is available in the group of Dr. Claudia Barros, Peninsula School of Medicine, Plymouth University, UK. The project will examine the role of novel candidates genes in brain tumour initiation and growth, following a single-cell transcriptome screen performed in the laboratory. Drosophila will be used to test candidate genes in the conversion of normal neural stem lineage cells into brain tumour initiation cells, via genetic gain and loss of function assays. Translation of findings into human-based systems will be performed by assessing human gene orthologues in glioma samples and patient-derived glioblastoma stem cell lines.
We are looking for a highly motivated graduate with a degree in biomedical Sciences or related field (1st class or 2:1 equivalent), and preferably with a relevant Masters qualification. International students must also have an IELTS score of 7.0 or above (or equivalent qualification). Experience in genetics, molecular biology, immunohistochemistry, cell culture, protein work and/or imaging is desired. A high interest in neural stem cells and brain tumour biology is essential.
Follow the link: https://www.plymouth.ac.uk/study/postgraduate and click ‘Apply’ to access the application form. Please mark the application for the attention of Bernice Wilmshurst and indicate the project Reference. Please Note: you do not need to submit a project proposal with your application.
Funding notes: The stipend will be £14,057 (based on full time 2015/16 rate). Tuition fees will be paid at the home/EU rate. Candidates who are not eligible for Home/EU fees will be liable for the difference between ‘home student fees’ and ‘international student fees’. For the 15/16 academic year the difference in fee is £10,800. If you are selected you will be required to provide financial assurances.
Closing date for applications: Noon 21st March 2016. Shortlisted candidates will be invited for interview. We regret that we may not be able to respond to all applications. Applicants that have not been contacted by 11th May 2016 should consider their application unsuccessful on this occasion.
In Spring 2015, just a couple of months into my PhD, I started to settle with my new surroundings in Zürich, making friends in my PhD lab of Dr. Christian Mosimann, and learning the fine details of early zebrafish development. That is when suddenly one morning Christian casually asked me how I’d feel about moving to Dresden in Germany for a while…
My key interest is the molecular control of cell migration and rearrangements. In our lab, we use the zebrafish (Danio rerio) to investigate the cell fate determination of the lateral plate mesoderm (LPM). The LPM is a fascinating mesoderm lineage that gives rise to the heart, blood, vessels, kidneys, and limbs. After gastrulation, the LPM in zebrafish arranges in tiers of cells at the lateral edge of the developing embryo. How the LPM originally arises from the remaining mesoderm, what common molecular program links its vast array of cell fates, and how early LPM cells arrange at their lateral positions remain unclear. I am particularly interested in the cell migration dynamics and migration control of the early LPM. To visualize the emerging LPM, the Mosimann lab has generated several novel transgenic zebrafish strains that mark the LPM cells at all stages of development. Nonetheless, traditional confocal imaging only enables imaging of small parts of the LPM and not of the whole embryo in its entirety.
A great solution to image the entire zebrafish embryo is selective plane illumination microscopy (SPIM), also called lightsheet microscopy. While we do have a local SPIM setup, processing the huge amounts of data accumulated is time-consuming. The lab of Jan Huisken, at the Max Planck Institute (MPI-CGB) in Dresden (Germany) is pioneering the development of high-speed fluorescence microscopy to enable systematic studying of developmental processes. Their selective plane illumination microscopy (SPIM) allows imaging several zebrafish embryos at the same time, overnight, and enables visualizing the whole zebrafish early development.
Gopi Shah, a senior grad student in the Huisken lab, had given a great presentation on panoramic SPIM at the European Zebrafish meeting in 2013, and Christian was immediately hooked. After several discussions between Jan and Christian and trading postdoc stories from their times at opposite coasts in the USA, Jan generously offered to have me come to his lab in Dresden and use their unique lightsheet setup to image our unique transgenic zebrafish lines. I was immediately enthusiastic about this great opportunity as I could learn a new technique, apply it to fascinating biology, and learn more about cell migration dynamics in vivo.
As the whole panoramic SPIM setup is too bulky to quickly ship to Switzerland, I moved for a month to Dresden. My arrival in the lab was warm and welcoming. Gopi introduced me into their lightsheet setup and supported me greatly with the imaging. Also the rest of the lab unconditionally helped me out during my stay. Although research with zebrafish is always an adventure, my zebrafish cooperated really well and I could gather more data than I had hoped for: I successfully managed to image over 60 zebrafish embryos.
Despite the many hours spend “behind” (or rather next to and around) the SPIM setup and on data processing, I also had the opportunity to fully explore beautiful Dresden, visit the Elbe Valley with its castles, and hike in the surrounding nature, especially in the national park referred to as Sächische Schweiz (Saxonian Switzerland).
Already just looking at the imaging provides us with new insights about the emerging LPM and early zebrafish development. For the first time, can we see the migration and organization of the whole LPM from its onset during gastrulation. This unique data set is a key fundament for my future studies of LPM patterning and for deciphering the cell migration dynamics in the emerging LPM. We have already lined up follow-up experiments coordinated between our both labs, and I continue to receive great help from the Huisken lab with analyzing all the data.
My collaborative visit was a definite success based on the amount of data I collected, the new ideas and immediate follow-up experiments, and the new contacts and friends that I made. It was a great experience to be in a lab focused on a different discipline, but with the same greater goal in mind: studying the molecular and cellular processes of early development. I am very grateful to Dr. Jan Huisken for giving me the possibility to use his lab’s infrastructure and hospitality, and to The Company of Biologists for the support in form of a Development Travelling Fellowship.
Aneuploid cells—that is to say those with an abnormal number of chromosomes—are found in most human tumours.
A study conducted at IRB Barcelona on the fly Drosophila reveals how surviving aneuploid cells favour tumour development.
Barcelona, Thursday 9th February 2016.- A recent analysis of 43,205 human tumours unveiled that 68% of solid tumours are aneuploid, that is to say, they have an altered number of chromosomes. In recent years, scientists have attempted to clarify whether this aneuploidy contributes to tumour development or whether it is a co-lateral effect of the genomic instability of cancer cells, which increase the rate of mutations and the likelihood of cancer.
A study by the group headed by ICREA researcher Marco Milán, at the Institute for Research in Biomedicine (IRB Barcelona), published in this week’s issue of Developmental Cell provides details of the relationship between genomic instability, aneuploidy, and cancer.
The study, which has also involved the collaboration of ICREA researcher Angel R. Nebreda, in the Oncology’s programme at the same institute, explains how the molecular and cellular mechanisms triggered by aneuploid cells can give rise to tumours.
The research on aneuploidy and tumorigenesis has been performed using the wing primordia of the fruit fly Drosophila melanogaster as a model. This tissue is an epithelium organised into a single layer and that grows by 20 to 30,000 cells in a few days. Given these features, this tissue is an ideal system in which to generate genomic instability and to dissect the cell and molecular mechanisms that elicit aneuploid cells in a proliferating tissue.
Aneuploid cells: first step, suicide
The team of researchers observed that aneuploid cells first activate apoptosis (or programmed cell suicide). At the same time, in an attempt to counteract the imminent loss of cells, they send signals to neighbouring ones instructing them to divide and proliferate further to ensure the development of normal tissue—in this case the fly wing. Next, they also activate a series of DNA repair signals and also anti-tumour protection in order to prevent further aneuploidy.
“We have described the cascade of cell and molecular processes, and repair defence and compensation mechanisms which, simultaneously or sequentially, are triggered in and by aneuploid cells,” explains the postdoctoral researcher Marta Clemente, first author of the study.
But what happens if aneuploid cells manage to survive? After preventing the cells from dying, the researchers observed that the proliferation signals derived from aneuploid cells, which previously served to maintain healthy tissue, now favoured tumour development.
This study widens the Darwinian perspective of genomic stability in the development of cancer, “perhaps an incomplete view of the role of genomic stability in tumorigenesis” says Milán. Such a perspective is based on a random increase in tumour-promoting genes and a loss of tumour-supressing genes, which ultimately favours the tumour cell.
“Somehow the aneuploidy derived from this genomic instability also causes metabolic stress, which in turn leads to the expression of a series of signals that can enhance tumour growth and development”.
Given that aneuploidy is common to most cancers, Marco Milán considers that searching for treatments directed exclusively at removing aneuploid cells may provide a good strategy to tackle them.
“This basic biology research provides new information about the molecular links triggered by aneuploid cells, and this is the step prior to studying possible therapies to combat cancer,” says the IRB Barcelona researcher.
Postdoctoral position to study development of the diaphragm muscle and congenital diaphragmatic hernias (CDH) using mouse genetics, in vivo and in vitro studies, 2 photon imaging of live embryos, and genomic data from CDH patients.
The diaphragm is the most essential mammalian skeletal muscle – vital for respiration and a critical barrier between the thoracic and abdominal cavities. Defects in diaphragm development are the cause of CDH – a common and often lethal birth defect. Despite the diaphragm’s functional importance and the frequency and severity of CDH, how the diaphragm develops and the etiology of CDH are only just beginning to be understood. The goals of this postdoctoral position are to build upon our recent research insights (Merrell et al. Nature Genetics 2015 and highlighted in the New York Times) and explore the genetic, molecular, and cellular mechanisms regulating the development of the diaphragm and determine how these processes are mis-regulated in CDH.
We are seeking a motivated, enthusiastic, and hard-working postdoctoral fellow to join our research team. The position is in the lab of Gabrielle Kardon (http://kardon.genetics.utah.edu/). The lab is located in the Department of Human Genetics at the University of Utah in Salt Lake City, providing a strong and collaborative research community, comfortable lifestyle, and ample opportunities for outdoor recreation.
Looking for postdoc to start anytime between Feb and Dec 2016. Please contact Gabrielle Kardon (gkardon@genetics.utah.edu) with CV, list of references, and a brief statement about why you are interested in the position. PhD in biology or biochemistry is required.
– And it seems that CRISPR will continue to make headlines in 2016. Just last week the lab of Kathy Niakan at the Crick Institute received the first licence to edit the genome of human embryos in the UK.
– Are biological databases like FlyBase, WormBase and ZFIN at risk of no longer being funded? Article in the Genes to Genome GSA blog discusses the issue.
– Science4Refugees is a new initiative to help refugee scientists and researchers find suitable employment in Europe.
– Why are certain science myths so enduring, and how are they holding back science?
– How scientists are helping movie writers make films ‘plausible-ish’- in The Wall Street Journal
– And the SDB has announced the winners of their 2016 Awards, including the SDB Lifetime Achievement Award to David McClay (Duke University) and the Conklin Medal to Kathryn Anderson (Sloan Kettering Institute). Meanwhile, the BSDB announced that the first Cheryll Tickle Medal will be awarded to Abigail Tucker (King’s College London).
Modern biology is impossible to understand without genetics. Students today would struggle to understand Mendelian inheritance without the idea of the gene, and labs worldwide use molecular biology and genetic techniques to study different organisms and processes. Genetics has revolutionised the study of biology in the past century, and the study of genetic material keeps providing insights into fields such as developmental biology and evolution. However, genetics has also brought with it many new words, from abstract concepts to names for processes or structures, and to the untrained, it can sometimes seem daunting to approach technical texts for lack of definitions. “Decoding the Language of Genetics”, by David Botstein, attempts to address this problem.
“Decoding the Language of Genetics” is a difficult book to categorise. It is not a textbook, in that it is readable, peppered with personal comments from the author, and at times avoids going into complex subjects in order not to confuse the reader, but it is also not a “popular science” book. It is very clearly an attempt to teach: not just to make science available, but to make it understood. Instead of running away from jargon, it attempts to explain it. In the introduction, David Botstein explains that he wanted to write a book that would allow the non-geneticist to understand the geneticists’ technical vocabulary, allowing them to access technical writing by geneticists. This implies that the book is not aimed at the general public, but rather intended for those with a reasonably robust knowledge of biology in general, but not of genetics specifically.
The book starts with a very valuable introduction to basic concepts in genetics (phenotype, genotype, etc.), but failing to define gene (Botstein describes the history of the word, but does not give a formal definition, possibly because he will later define locus and functional gene, and at that early stage the preliminary concepts needed to define gene have not yet been presented). Botstein then proceeds to define commonly used acronyms in modern genetics and explain the experimental origin of such acronyms. The book proceeds in this fashion, explaining not just the vocabulary, but also why it is needed (and in some cases, why it is not). As it continues, the book goes from explaining specific concepts to explaining complex experiments and the effects they have had in both the science of genetics and in its vocabulary. Botstein often gives reasons as to why certain words are used over others, and occasionally explains his own preference for certain expressions, usually based on their clarity or specificity. As an example, he favours using synthetic phenotype over epistasis when talking about the phenotypic consequences of interactions between several genes simply because epistasis has been used very differently by different people, and he has found that the word can be confusing for his students.
The book excels where it explains concepts and supports their explanation with clear descriptions of the experiments performed to arrive at those concepts. A particularly enjoyable example of this is the chapter on functional suppression (rescue of a mutant phenotype via a mutation in a second gene that compensates for the original mutation). Botstein first gives a general definition of the concept, comparing it to previously described informational suppression. He then proceeds to name different types of functional suppressors, giving specific examples and explaining how they were discovered. He uses Georgopoulos’ and Herskowitz’s experiments with λ phage and Escherichia coli to illustrate ; Jarvik’s and Moir’s experiments with heat and cold sensitive mutants to talk about suppressors with novel phenotypes; Hodgkin’s experiments on sex determination in to describe recessive functional suppressors; and yet others’ experiments to make bypass suppression, dosage suppression and mutual interaction suppression understood. It is not surprising, therefore, that the books’ weakest points are when the author fails to give examples to illustrate a concept, or to fully explain why something occurs. This happens specially in the chapter about recombination and linkage mapping, where (to give one example) Botstein writes about the frequency of recombination: “Frec= (number of recombinant gametes)/(total gametes). This parameter can vary from 0 to 0.5”. However, he fails to give an explanation as to why the maximum is 0.5 and not 1. It may be that Botstein thought his readership would know that that these numbers are the result of an experimental observation, but having explained simpler concepts earlier in the book, it was surprising he did not provide a short sentence making this clear.
“Decoding the language of genetics” is laid out in a hierarchical fashion: starting off with basic definitions, it moves on to concepts of increasing complexity. Although this is a fantastic approach, since it allows the less knowledgeable reader to build on their knowledge, an academic audience may find the first few chapters rather basic, and give up on the book without realising that the explanations provided are necessary to understand the rest of the work. This approach also means that Botstein occasionally fails to give a definition early in the text (even though he is using the word and discussing it’s meaning, as in the case of “gene”) in favour of giving a more complete one at a later point, and this can make the reader feel that definitions are missing and, therefore, that the book is failing to do what it claims.
When I started reading “Decoding the Language of Genetics”, I didn’t understand who it was meant for very well. I found the concepts described too basic for the academic reader, but the tone too academic for the lay person. As I continued, I started to realise I was learning new things, and by the end of the book I came to a conclusion. It is the perfect book for any bioscience researcher who will encounter genetics vocabulary in their work, but who hasn’t had a thorough training in genetics. Some of the concepts at the beginning will seem basic, because the researcher will almost certainly have studied genetics at some point, but they are definitions that need to be made explicit for the concepts to be used later on. The emphasis on detailing how classical genetics experiments were performed, and how conclusions about gene and genome structure were extracted from these experiments, becomes a fantastic lesson, not only in the history of genetics, but also in experimental design and interpretation.
This is a review of “Decoding the Language of Genetics“, by David Botstein. Published by Cold Spring Harbor Laboratory Press in 2015. ISBN: 978-1-621820-92-5.
Hi everyone! I’m Helena. Some of you may know me as the current intern here at the Node, but next week, I will go back to Alfonso Martínez Arias’ lab at the University of Cambridge to continue working on my PhD. Our lab is interested in cell fate and differentiation in the context of early development. In particular, we want to understand how interactions between signals and gene regulatory networks can generate tissues and organs from single cells.
In order to study all of these phenomena, we use embryonic stem cells (ESCs). Specifically, in my project I use mouse embryonic stem cells (mESCs). mESCs are an incredibly useful system to study both cell fate decisions and early development because they are relatively easy to maintain in vitro and they are easy to access and observe; much easier than studying early development in vivo where the embryo is not always so easily accessible. Since mESCs are grown in culture, it is possible to achieve a very stringent control over the inputs the cells receive: I can control the medium the cells are grown in, the temperature, the atmosphere, what signals are applied to them and when, etc. You’d think that with so much control, I would know exactly what to expect every time I look into a tissue culture flask, but ESCs can be extremely finicky: one tiny change, for example a different batch of media or a slightly different density of cells can turn a good culture bad and throw weeks of work down the drain. Not to mention that different ESC lines all grow better in slightly different conditions, meaning each time I start using a new line, or when I culture several lines at a time, I have to adapt. However, one of the major cell drawbacks to cell culture when studying development is that the structural and mechanical constraints of the embryo are missing. Despite these limitations, mESCs are a very powerful model to study how cells respond to their environment and the types of inputs that result in maintenance of or exit from pluripotency and differentiation to specific lineages.
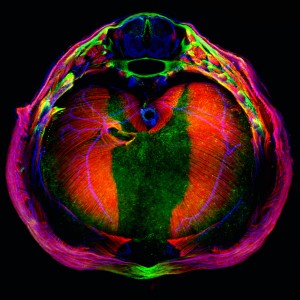
Mouse embryonic stem cell colony stained for Oct4 (in cyan). Oct4 is a transcription factor expressed in pluripotent cells, and it has roles in both the maintenance and the exit of pluripotency.
A typical day in an ESC lab varies considerably depending on whether one is running experiments or not, and also on the stage of the experiment. Since people in the lab are interested in how signals and gene networks interact, and because ESC culture is a model system that lends itself to producing large datasets (a single experiment can involve millions of mESCs, so potentially, millions of observations), the lab also does a lot of statistical analysis and modelling, alongside experiments. There are certain everyday tasks, however, that are fairly consistent throughout most stem cell labs, in particular tissue culture.
ESCs are fussy creatures, and require daily attention in order to maintain them in good condition in culture. This is because the function of ESCs in vivo is to divide, differentiate and build an embryo, so unless one grows them under certain conditions that promote pluripotency (i.e., with inhibitors of Wnt and FGF signalling), they are naturally inclined to differentiate. ESCs can be grown with or without feeder cells (a layer of cells underneath the ESCs that support growth, usually embryonic fibroblasts) and with our without antibiotics (to prevent infection). In our lab, we usually don’t use either: feeder cells can complicate both experiments and data analysis (for example in the case of RNA-seq experiments), and antibiotics can mask infections affecting growth and leading to contaminated experiments. ESCs are grown in flasks with cell medium, which needs to be changed every day. A lot of care has to be taken to avoid contamination: bacteria will happily grow in the nutrient-rich mESC media, eventually killing the cells. When we do have the occasional outbreak, tensions run high in the lab: a contaminated experiment is a useless one, and once a culture is contaminated, it’s not unusual for others to become infected. It also means that the tissue culture cabinets have to be meticulously cleaned with fungicide, ethanol and UV treatment – a job that no one is ever keen to take on. In our lab, we have to periodically remind each other of this, with varying degrees of severity in our reminders.
In addition to daily media changes, mESCs also need to be passaged every so often. In my case, I usually passage cells every two days, but this depends on the density of the cells in the flask. Passaging (or splitting) cells means transferring a proportion of the cells into a new flask. This is necessary in order to make sure the culture stays at the correct density, rather than becoming overgrown. This helps to maintain pluripotency and ensures that the cells are in optimal condition to perform experiments. mESCs are grown in adherent culture, meaning that the cells are attached to a matrix present on the surface of the culture flask – in our lab we use gelatin. The first thing we do when we passage cells is to apply trypsin to detach the cells from the surface of the flask. Then, we add medium to neutralise the trypsin and to dilute the cells. At this point, some people prefer to collect all the medium, spin down the cells and resuspend them before transferring into a new flask, in order to get a more accurate passage. I usually do this when I’m performing an experiment, or when I’m using a new cell line to be more accurate, but in the day to day, I will often skip this step and simply take a portion of the media I’ve used to neutralise the trypsin and transfer it to a new flask.
It is always fun to see someone new learning tissue culture. I’ve seen people with their whole heads inside a cabinet to avoid hitting the glass while working! This is obviously not the best approach for sterile technique. On one occasion, I even saw someone spray their cells with ethanol to avoid contamination. In all fairness, they achieved their goal of not contaminating their culture, although their cells died during the process! Learning tissue culture is mostly about learning good habits, although individual habits can vary from person to person. For example, in my lab, the direction pipettes are placed in the hood can be a point of contention between different people. The tissue culture cabinets can get pretty busy at times, but we all know (for the most part) when others like to do their tissue culture work, which helps with scheduling so we can all get our work done.
Once the cells have been taken care of, there are still plenty of things to do in the lab. mESC experiments usually involve growing cells in different conditions and analysing them at specific intervals. In our lab, the goal of experiments is usually to determine how the conditions used have affected the state of the cells. Has the cell exited pluripotency? If it has and it is differentiating, what is it differentiating into? How does this relate to what happens in the embryo? In order to answer these questions, we perform immunohistochemistry experiments, tracking experiments (time-lapse imaging of cell lines with fluorescent reporters for the expression of a gene of interest), flow cytometry analyses, RNA-seq, Western blots, etc. In addition, people in ESC labs spend considerable amounts of time generating new cell lines. These can be anything from cells with fluorescent reporters for the expression of genes of interest to knockout lines. As you have probably guessed, as a stem cell scientist you have to be in the lab every day when you are culturing cells. Sometimes I miss being able to go away for a weekend on a whim, but for the most part, I enjoy the flexibility. Working on the weekends can have its upsides: it means I’m not constrained by five-day experiments, and I can also use facilities that are usually busy during the week! Of course, everyone in the lab wants a weekend off once in a while, so we often “babysit” each other’s cells for a weekend. The nicer side to tissue culture is that, even though you have to do it every day, there will be days when the only thing to do is to change media – a process that can take as little as five minutes. With that done, you can spend the rest of the day catching up on reading, analysing data, imaging, and of course, on extended coffee/tea breaks chatting with your colleagues – about science of course!
This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series here and read other posts in this series here.
We have an offering an exciting post-doctoral opportunity to image and quantitatively analyse tiny transparent ascidian embryos with a brand new multi view light sheet microscope. Apply now!

 (No Ratings Yet)
(No Ratings Yet) (1 votes)
(1 votes)


![[Parameter-Settings] FileVersion= 2000 Date/Time = 0000:00:00 00:00:00 Date/Time + ms = 0000:00:00,00:00:00:000 User Name = TCS User Width = 1024 Length = 1024 Bits per Sample = 8 Used Bits per Sample = 8 Samples per Pixel = 3 ScanMode = xy Series Name = ptc_msl1p35_MMP1 p35 4d29.lei](https://thenode.biologists.com/wp-content/uploads/2016/02/DrosWing_LBarrio1-300x300.jpg)


 This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series
This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series 