A regeneration retrospective: planarians aplenty
Posted by Alex Eve, on 3 September 2023
This post is part of the regeneration retrospective series.
In some ways, it’s difficult to imagine a better model of regeneration than the planarian flatworm. These comical, googly-eyed creatures have captivated scientists for centuries with their remarkable regenerative potential (Dalyell., 1814; Abeloos, 1930). Famously, planarians can be cut into small fragments, each of which is capable of regenerating the full body, (including the brain, eye-spots and intestine), an ability owed to the distribution of potent adult stem cells, known as neoblasts, which are distributed throughout the body and ‘hyperproliferate’ following amputation (Ivankovic et al., 2019; Lee et a., 2022). In this first instalment, I discuss the highlights from the following two articles:
Size of Fragment and Rate of Regeneration in Planarians
Agnes Brøndsted and H. V. Brøndsted
https://doi.org/10.1242/dev.2.1.49
Danish couple, Agnes and Holger Brøndsted, married in 1919 and had the culmination of their work studying planarian fragment size regeneration published in March 1954, in the first issue of the second volume of JEEM/Development (Brøndsted and Brøndsted, 1954). At this time, the regenerative ability of planarian fragments was well established and the presence of neoblasts was accepted. However, Agnes and Holger’s article came at a time of confusion in the field, when conflicting evidence made it unclear whether the number of cells in each fragment contributed to the speed at which the fragment regenerated. They set out to address this question with the minimalistic approach of cutting animals into fragments of different sizes and measuring the length of time eye-spots took to regenerate. Their work began by circling the shores of Lake Fures near Copenhagen, Denmark, lifting rocks to collect planarians from the wild. They ended up with a collection of a few hundred animals from four different species: Dendrocoelum lacteum, Bdellocephala punctata, Euplanaria lugubris and Polycelis nigra. The authors highlight that these four species do not reproduce asexually by fission, removing a confounding variable from their studies. For reasons, I admit, aren’t so clear to me, they dissect each of the species in different ways to produce fragments of different sizes (Fig. 1). However, regardless of the size or shape of the fragment, the rate of regeneration was the same, showing conclusively that the number of cells (and thus the number of neoblasts) in the fragment does not contribute to regeneration speed, suggesting that neoblasts are unlike to migrate within the fragment.

Around 70 years later, planarians are still a potent and popular organism for studying regeneration; however, the variety of species used as models has largely converged on two species: Schmidtea mediterranea and Dugesia japonica. Significant technical advances, not least of which the culture of planaria in the lab, have allowed researchers to understand the cellular and molecular mechanisms of stem cell behaviour during regeneration and mean that hunting for critters under rocks is now a pastime, rather than a scientific necessity.
Loss of plac8 expression rapidly leads pluripotent stem cells to enter active state during planarian regeneration
Hayoung Lee, Kanon Hikasa, Yoshihiko Umesono, Tetsutaro Hayashi, Kiyokazu Agata and Norito Shibata
https://doi.org/10.1242/dev.199449
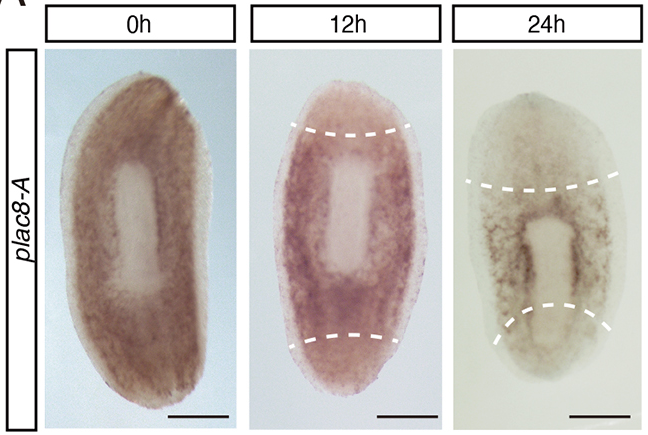
Last year in Development, Hayoung Lee (Kyoto University) and colleagues capitalised on modern innovations, such as markers for neoblasts and genetic knockdown experiments, to understand the role of a specific gene, plac8-A, during regeneration in D. japonica (Lee et al., 2022). Unlike the species used by the Brøndsteds, D. japonica can reproduce asexually via fission. The authors of the present study employ this fission phenomenon as a measure of cell proliferation and show that knockdown of plac8-A increases the number of fission events. Using imaging and expression analyses, Lee and colleagues demonstrate that plac8-A is expressed by neoblasts and is downregulated before the neoblasts proliferate (Fig. 2). Finally, through chemical treatments, Lee and colleagues show that ERK signalling acts upstream of plac8-A and inhibits plac8-A expression during regeneration via the JNK signalling pathway. Together, these findings demonstrate that plac8-A acts as a molecular switch that regulates neoblasts’ regenerative behaviour.
Together, these two articles address a similar question: What influences neoblast proliferation during planarian regeneration? The Brøndsteds show that, following amputation, not all cells hyperproliferate and that the quantity of cells doesn’t regulate this process, while Lee and colleagues determine that plac8-A must be downregulated in order for neoblast hyperproliferation to occur and loss of plac8-A can induce neoblast proliferation across the whole organism. I hope you enjoyed the post and join me again tomorrow when we will be revisiting the archives to look back on skeletal muscle regeneration.
References
M. Abeloos. Recherches expérimentales sur la croissance et la régénération chez les Planaires. Bull. biol. 1930; 64, 1–140.
Agnes Brøndsted, H. V. Brøndsted; Size of Fragment and Rate of Regeneration in Planarians. Development 1 March 1954; 2 (1): 49–54. doi: https://doi.org/10.1242/dev.2.1.49
J. G. Dalyell. Observations on Some Interesting Phenomena in Animal Physiology, Exhibited by Several Species of Planariae: Illustrated by Coloured Figures Of Living Animals. 1814. Edinburgh: Archibald Constable & Co.
Mario Ivankovic, Radmila Haneckova, Albert Thommen, Markus A. Grohme, Miquel Vila-Farré, Steffen Werner, Jochen C. Rink; Model systems for regeneration: planarians. Development 1 September 2019; 146 (17): dev167684. doi: https://doi.org/10.1242/dev.167684
Hayoung Lee, Kanon Hikasa, Yoshihiko Umesono, Tetsutaro Hayashi, Kiyokazu Agata, Norito Shibata; Loss of plac8 expression rapidly leads pluripotent stem cells to enter active state during planarian regeneration. Development 1 February 2022; 149 (3): dev199449. doi: https://doi.org/10.1242/dev.199449
 (No Ratings Yet)
(No Ratings Yet)
 (1 votes)
(1 votes)