A PhD project proposal to be carried out in Patrick Lemaire’s lab at CRBM, Montpellier, France, in tight collaboration with Grégoire Malandain, Morpheme INRIA team, Sophia-Antipolis, France. Funding is conditional upon selection by the CBS2 doctoral school‘s entrance jury.
Why study ascidians?
The embryonic development of ascidians, a group of marine invertebrates, is remarkably conserved, at the single cell level, between individuals of a given species and between species, even if they diverged up to 400 MY ago. Ascidian genomes, however, evolve particularly rapidly. The remarkably simple and transparent ascidian embryos are thus ideal to study developmental systems drift and to identify constraints that could explain the exceptional evolutionary precision and stability of embryonic morphologies.
The proposed project
We propose a computational PhD project, which will use experimental data collected in the team or in public databases to characterise inter-individual and inter-species variability at the geometric, mechanical and transcriptional scales.
This project will involve the development of concepts and computer tools to study and measure, at each scale studied, the variability of different parameters (variability of cell lineages, cell lifetimes, orientation of cell division, pressures, surface and line tensions, gene expression, etc.). These studies will lead to a reflection on the concept of the average embryo and on its computational representation. The tools developed will open the way to the quantitative study of robustness to environmental and genetic perturbations, and to the identification of bridges between scales of analysis (search for co-varying parameters across scales).
What is available to start the project?
The project will benefit from the conceptual and methodological developments made over the last 10 years by the hosting teams and their collaborators (see references below). These breakthroughs (ASTEC, MorphoNet, Aniseed) place them in a unique position to analyse, experimentally and computationally, the variability of animal embryogenesis, whether natural or in response to environmental or experimental perturbations.
Using these advanced tools, we generated high resolution geometric and mechanical descriptions of 7 embryos of the ascidian Phallusia mammillata, over several hours of development and with a 2 minutes time resolution. You can view a short video highlighting this works.
These embryos and tools constitute a solid basis for the proposed PhD project. The embryo collection is currently expanding to include more WT Phallusia embryos as well as embryos cultured in response to environmental (temperature, salinity, pH) or genetic perturbations.
How to apply?
The ideal candidate will have successfully graduated from a Master’s programme in computer science, bioinformatics or physics recognized by France. S/He will have strong computational skills and some knowledge of developmental biology. A working knowledge of English (B2) is needed. There is no prerequisite in French.
To apply to the project, please contact P. Lemaire (patrick.lemaire[at]crbm.cnrs.fr) as soon as possible and by May 11, 2022 at the latest with a motivation letter, a CV and the names and contact details of 2 academic referees including the PhD supervisor.
References
Guignard L. *, Fiuza U.-M. *, Leggio B., Laussu J., Faure E., Michelin G., Biasuz K., Hufnagel L., Malandain G. #, Godin C. #, Lemaire P.# (2020) Contact-area dependent cell communications and the morphological invariance of ascidian embryogenesis. Science, 369 :6500 eaar5663
Dardaillon, J; Dauga, D; …; Dantec, C.#; Lemaire, P#. (2019) ANISEED 2019: 4D exploration of genetic data for an extended range of tunicates. Nucleic Acids Res. 48(D1): D668-D675
Leggio, B; Laussu J; Carlier, A; Godin, C; Lemaire, P and Faure, E (2019) MorphoNet: An interactive online morphological browser to explore complex multi-scale data. Nat Commun. 10(1):2812
The tunicate team at CRBM (Montpellier, France), headed by Patrick Lemaire, is offering a 2-year post-doctoral fellowship (or a 3-year PhD fellowship for an exceptional candidate) to study the robustness of animal embryonic development to genetic and environmental perturbations, using quantitative live imaging of ascidian embryos. A short video describes a recent piece of work of the team relevant to the project.
Why ascidians?
Ascidians are a group of marine invertebrates. Their embryonic cell lineages and early embryonic stage morphologies have remained essentially identical since the group’s emergence about 400 million years ago. This suggests that they are subject to very strong developmental or evolutionary constraints (Lemaire et al. 2011). Ascidian embryogenesis is also very robust to environmental perturbations of temperature and salinity. The extreme evolutionary and environmental robustness of embryonic geometries and cell lineages contrasts with a rapid genetic divergence between species and intra- and inter-specific variability in gene expression.
What is the project?
The proposed experimental project will provide a quantitative assessment of the developmental robustness of a critical morphogenetic process, ascidian gastrulation (1, 3, 5), to two key environmental parameters (water temperature and salinity) and to genetic perturbations of the morphogenetic driving force apparatus (myosin II, Rho kinase, …). The project will study the magnitude of environmental or genetic variations compatible with the production of a viable larva. It will seek to identify the least – and most – robust developmental processes and time points, i. e. those that collapse first, or resist best to the perturbations. Finally, it will characterize the structure of the natural and experimentally-induced variability in the geometry and mechanical properties of embryos. This may lead to the identification of developmental modules.
The project will involve advanced light-sheet imaging of live micro-injected embryos of the ascidian Phallusia mammillata, followed by the computational and statistical analysis of the acquired developmental movies (see 2, 4).
Who funds the project, and how to apply?
The project is funded by an ANR-NSF binational project grant and will be conducted in collaboration with 3 other teams: Prof. Atef Asnacios (MSC, Paris), Prof. Edwin Munro (U. Chicago, USA) and Prof. Madhav Mani (Northwestern University, Evanston, USA).
Expected candidates will have a PhD in cell and developmental biology, an excellent track record of publications and oral communications, strong skills in fluorescent live imaging and some experience in the computational analysis of large datasets. To apply, send Patrick Lemaire (patrick.lemaire[at]crbm.cnrs.fr) by May 11, 2022 at the latest a motivation letter, a CV and the names and contact details of 2 academic referees including the PhD supervisor. A working knowledge of English (B2) is needed, there is no prerequisite in French.
References
Fiuza U.-M. and Lemaire, P. (2021) Mechanical and genetic control of ascidian endoderm invagination during gastrulation, Semin Cell Dev Biol 120:108-118
Guignard L. *, Fiuza U.-M. *, Leggio B., Laussu J., Faure E., Michelin G., Biasuz K., Hufnagel L., Malandain G. #, Godin C. #, Lemaire P.# (2020) Contact-area dependent cell communications and the morphological invariance of ascidian embryogenesis. Science, 369 :6500 eaar5663
Fiuza U-M, Negishi T., Rouan A., Yasuo H.#, Lemaire P. # (2020) Nodal and Eph signalling relay drives the transition between apical constriction and apico-basal shortening during ascidian endoderm invagination. Development 147: dev186965
Leggio, B; Laussu J; Carlier, A; Godin, C; Lemaire, P and Faure, E (2019) MorphoNet: An interactive online morphological browser to explore complex multi-scale data. Nat Commun.10(1):2812
Sherrard, K., Robin, FB, Lemaire, P., and Munro, E. (2010) Sequential activation of apical and basolateral myosin drives endoderm invagination during ascidian gastrulation, Current Biology, 20(17):1499-510.
Read on for our news roundup of the past month, with an emphasis on what has caught our eyes on twitter. This month, we also include a list of meetings with upcoming deadlines, as well as a selection of preLights.
#leavingacademia
At the beginning of the month, it seemed like everyone was #leavingacademia. Below are a few of our favourite threads that highlight some of the challenges that academics face, but also the upsides of academic research and working with industry.
1/9. There has been a ton of discussion on academic Twitter about reasons to leave academia. This thread is for young scientists who want to stay in academia. Let me first comment on the things I agree with from the “time to leave” tweets:
(deep sigh) I think the current debate about academia vs. industry does not articulate how it really is better for all of us that there is robust competition and introspection about what job opportunities are present to us, and more importantly to our trainees.
It has been great to see the return of so many in-person conference. With ‘conference fever’ in mind, we have put together a list of #devbio and #stemcell meeting with upcoming abstract and registration deadlines. You can find a list of upcoming conferences on our events page. If we are missing your event, then please feel free to add it to our planner.
If you are interested in science communication, would like to improve your writing skills and become part of the preLights community, don’t miss the open call for new preLighters.
If you would like to write for the Node, check out our recent list of writing ideas. If you would like to contribute to our ‘Developing news’ blog, please get in touch at thenode@biologists.com
My journey in the field of mammalian developmental biology began when I joined the lab of Dr Ivan Bedzhov at the Max Planck Institute for Molecular Biomedicine (MPI-MB) in 2016 to pursue my doctoral research. After Ivan and I discussed the many important processes that take place during the peri-implantation phase of embryonic development, it was the unaddressed mechanisms by which the embryo forms the first contacts with the mother during implantation that caught my interest the most.
The process of implantation mediates the first direct interactions between the embryo and the mother. Specialized cells of the mouse embryo, known as trophoblast giant cells (TGCs), invade deep into the uterine tissues, enabling nutrient uptake and gas exchange with the maternal environment. In turn, the uterine stroma rapidly proliferates and completely engulfs the implanting embryo. Thus, studying the process of implantation is fundamentally challenging, as the embryo is concealed by the uterine tissues and its development depends on the maternal support.
According to clinical reports, almost half of the human pregnancies fail at the time of implantation (Boomsma et al., 2009; Koot et al., 2012). However, the cellular mechanisms of implantation and the factors causing termination of pregnancy are very poorly understood. The significance of these open questions and the excitement to understand the first interactions between an embryo and the mother motivated me to work on this project.
A glimpse at the trophoblast – maternal vasculature interactions in utero
As an early miscarriage can happen because of poor trophoblast penetration and/or inadequate blood supply to the implantation site (Klauber et al., 1997; Reus et al., 2013; Torry et al., 2007), we were keen to understand whether an active crosstalk between the TGCs and the maternal blood vessels takes place during the implantation stages. We examined the spatial organization of the TGCs and determined that the invasive trophoblast is organised as strands of cells penetrating the uterine stroma. Moreover, we found that the TGCs of the implanting embryo intermingle with the surrounding blood vessels (Fig. 1). As static images were insufficient to study the dynamics of these interactions, we decided to establish a 3D biomimetic platform that resembles the biomechanical properties of the uterine stroma.
Figure 1: Whole mount image of uteri containing E4.75 LifeAct GFP Embryo stained for – LifeAct GFP Embryo (GFP) in green, blood vessels (PECAM1) in red and nuclei (DAPI) in blue. White arrow indicates invasive trophoblast intermingling with the surrounding blood vessel.
Establishment of the 3D biomimetic platform
First, we determined the stiffness of the maternal tissues using atomic force microscopy (AFM) and examined the cell adhesive properties of the endometrium. As we also had to take into account that the TGCs secrete matrix metalloproteinases (Zhu et al., 2012), we used biodegradable synthetic hydrogels as our artificial substrate. We tuned the stiffness and adhesive features of the hydrogels to mimic the biomechanical properties of the uterine stroma, (in collaboration with Dr Britta Trappmann’s and Dr Adrian Ranga’s labs). Culturing embryos in this 3D environment enabled us to directly observe ex utero implantation of the embryos for the first time, which was very exciting!
As our main focus was to understand the crosstalk between the invading TGCs and the maternal vasculature, we teamed up with Hongyan Long from Dr Britta Trappmann’s lab to incorporate the implanting embryos into a microfluidic device (Trappmann et al., 2017) which allowed us to model the interactions with the blood vessels. After several months of tireless efforts and optimization, we managed to establish the right conditions for co-culturing embryos and endothelial cells in the microfluidic chip. Using this platform, we found that the invasive trophoblast migrates towards the blood vessels to form direct cell-cell contacts, similar to the static images in vivo (Movie 1). This was truly fascinating and a very rewarding moment for us.
Movie 1: Timelapse movie of LifeAct GFP embryo and tdTomato expressing endothelial cells co-cultured in microfluidic device
Uncovering the molecular basis of the trophoblast – vasculature interactions
Next, we wanted to examine the signalling crosstalk that mediates these interactions. We are grateful to Dr Hyun-Woo Jeong from Professor Dr Ralf Adams lab, who helped us with the bioinformatic analysis. We found that the TGCs gain expression of cell surface receptors, ligands and adhesion molecules, similar to the ones expressed in the nearby blood vessels. By functionally examining the main players in this process, we found that platelet-derived growth factor receptor (PDGFR) signalling promotes the establishment of direct cell-cell contacts between the TGCs and the vasculature. As PDGFR signalling has been previously shown to mediate the recruitment of pericytes to the endothelial cells (Lindahl et al., 1997), our findings suggested that the TGCs exploit this signalling pathway to locate the endothelial cells of the maternal blood vessels. Consequently, the expression of compatible cell adhesion molecules in the TGCs and the vasculature, such as VE-cadherin, enables the formation of heterologous cell-cell contacts. This discovery also has a potential clinical relevance, as it has been shown that the PDGFR inhibitor used for the treatment of chronic myeloid leukaemia increases the risk of miscarriage in experimental animals and pregnant patients (Salem et al., 2019; Ault et al., 2006; Pye et al., 2008) and the reason for this side effect was so far obscure.
Bumps along the way
Like most of the projects in academic research, the road of our journey to understand the embryo maternal interaction was not always smooth. We did face unfortunate situations, but at the same time we also got a lot of positive comments from colleagues around the world about the novelty of the project and this encouraged us that we were on the right path. Most of the methods used in this study required intense optimization. We did get stuck sometimes but we kept trying and found new ways to overcome technical limitations, which also helped me to expand my skills and critical thinking. I am really thankful to all our collaborators for their time to discuss and troubleshoot challenging experimental approaches. We learned a lot through failed experiments and eventually when we look at the new discoveries that we’ve made, I think it was all worth the effort!
When we submitted our work to two of the top journals, we experienced major delays as reviewers that accepted the invitation to review our manuscript did not respond and kept us waiting for several months. At this point we had no choice but to move to another journal, as we were running out of time and funding. This happened twice and we lost 9 months just waiting for the initial review, despite the best efforts of the editors of these journals. Having to wait so long was very stressful. Unfortunately, other colleagues have also had similar experiences, so it is up to us, the research community as well as the journals to prevent such toxic behaviour. We finally found a good home for our manuscript in Developmental Cell, where the editors are doing their best to ensure fair peer-review process and present exciting new research to the community.
In the End…
Looking back, this journey has taught me that passion, compassion, clear objectives, perseverance, and interdisciplinary collaborations are key for making new discoveries. Looking forward, I hope that one day, our findings will help scientists understand how the human embryo establishes its first direct contacts with the mother and will lead to therapeutic approaches that substantially decrease the risk of an early miscarriage.
Ault, P., Kantarjian, H., O’Brien, S., Faderl, S., Beran, M., Rios, M.B., Koller, C., Giles, F., Keating, M., Talpaz, M., et al. (2006). Pregnancy among patients with chronic myeloid leukemia treated with imatinib. J Clin Oncol 24, 1204-1208.
Boomsma, C.M., Kavelaars, A., Eijkemans, M.J., Lentjes, E.G., Fauser, B.C., Heijnen, C.J., and Macklon, N.S. (2009). Endometrial secretion analysis identifies a cytokine profile predictive of pregnancy in IVF. Hum Reprod 24, 1427-1435.
Klauber, N., Rohan, R.M., Flynn, E., and D’Amato, R.J. (1997). Critical components of the female reproductive pathway are suppressed by the angiogenesis inhibitor AGM-1470. Nat Med 3, 443-446.
Lindahl, P., Johansson, B.R., Leveen, P., and Betsholtz, C. (1997). Pericyte loss and microaneurysm formation in PDGF-B-deficient mice. Science 277, 242-245.
Pye, S.M., Cortes, J., Ault, P., Hatfield, A., Kantarjian, H., Pilot, R., Rosti, G., and Apperley, J.F. (2008). The effects of imatinib on pregnancy outcome. Blood 111, 5505-5508.
Reus, A.D., El-Harbachi, H., Rousian, M., Willemsen, S.P., Steegers-Theunissen, R.P.M., Steegers, E.A.P., and Exalto, N. (2013). Early first-trimester trophoblast volume in pregnancies that result in live birth or miscarriage. Ultrasound Obst Gyn 42, 577-584.
Salem, W., Li, K., Krapp, C., Ingles, S.A., Bartolomei, M.S., Chung, K., Paulson, R.J., Nowak, R.A., and McGinnis, L.K. (2019). Imatinib treatments have long-term impact on placentation and embryo survival. Sci Rep 9, 2535.
Torry, D.S., Leavenworth, J., Chang, M., Maheshwari, V., Groesch, K., Ball, E.R., and Torry, R.J. (2007). Angiogenesis in implantation. J Assist Reprod Genet 24, 303-315.
Our sister site, FocalPlane, has recently launched the FocalPlane Network, a global directory for microscopists. Like the Node Network, the FocalPlane Network was created with diversity in mind, and is entirely inclusive. Anyone with expertise in microscopy, including developers, imaging scientists, biologists and bioimage analysts, at any career stage, can enter themselves into the database. Members are also invited to provide diversity information such as gender, race/ethnicity, LGBTQ+ identity and disability status, although there is no requirement to do so. Users can access the network to search for potential collaborators, speakers, committee members and reviewers, using filters based on scientific expertise, geographic location, and aspects of diversity.
We would encourage anyone using microscopy in their research to add themselves to the database, and invite early-career researchers in particular who may be less visible to the microscopy community. Visit the FocalPlane site for more information about the network and FAQs.
“GENE AND CELL THERAPIES FOR HEALTHY AGEING” (virtual event)
April 12th 2022 (1-4 pm GMT, Lisbon Time),
PROGRAM: 13:00 Welcome – 13:05 Keynote: Bruno Silva-Santos, Instituto de Medicina Molecular João Lobo Antunes, Portugal. “Delta One T cells: development of a novel adoptive cell therapy for cancer” – 14:05 Clévio Nóbrega, Algarve Biomedical Research Institute, Faro, Portugal. “Towards modifying gene therapies for spinocerebellar ataxias” – 14:55 Break – 15:00 Miguel Sena-Esteves, UMass Chan Medical School, USA. “AAV gene therapy for neurological diseases” – 15:50 Closing Remarks
Welcome to the spring edition of our ‘Occasional writing ideas’ newsletter. With the world opening up again, we have focussed our ideas on getting back to ‘normal’ or finding our new normal. However, we are not limiting our posts to these topics, and you are welcome to post on any subject relevant to the developmental and stem cell biology community. If you would like to discuss any ideas for new content, or would like help with editing or posting, please get in contact at thenode@biologists.com.
Helen Zenner Community Manager, the Node The Company of Biologists
Writing Ideas
Meetings or symposia
Are you about to attend your first in-person meeting, or just your first for two years? You could share your experiences in a meeting report. See our blog on writing meeting reports here for guidance on what you can include in your post.
Will you be taking advantage of the fact that so many meetings now have a virtual component? Is this for public health, accessibility or sustainability reasons? What do you think conference providers can do to improve your experience?
Webinar series/online communities
Are you involved with a webinar series that you would like to highlight? Do you have any tips and tricks to overcome Zoom fatigue and keep your audience engaged?
Did you start, or become part of, an online community during the pandemic? How has this helped your mental health, your research, or your career?
Lab life
What are the positive and negative effects of the pandemic on your research? Has the pandemic led to any behavioural changes that are set to stay?
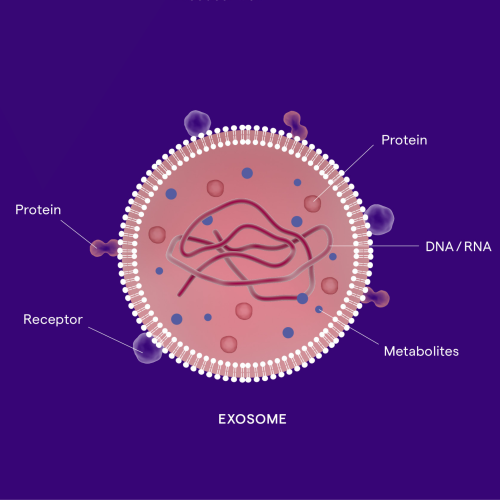
“We’ve tamed the toxicity of this cancer drug by attaching it to an exosome. Previously the side effects prevented it from becoming a treatment, but because it’s associated with an exosome, it stays in the lesion and we get all of the drug effects locally without the tolerability issues”
Dr Doug Williams, CEO of Codiak Biosciences
In the latest episode of the Genetics Unzipped podcast, sponsored by Lonza, unpacking the science behind exosomes: one of the hottest new areas of research for both diagnosing and treating diseases. Once thought to be little more than ‘dust’, exosomes are tiny biological mailbags that travel around the body, as Kat Arney hears from Rossella Crescitelli. Sally Le Page talks to Doug Williams to learn how exosomes can be used to reduce the toxicity of cancer treatments by targeting drugs directly to tumours. And Davide Zocco tells us how they are scaling up manufacturing to take exosome treatments from the lab to clinics.
In our latest SciArt profile, we meet Maria Abou Chakra. Maria is a theoretical biologist who introduces us to her sci-sketchnotes and highlights the importance of creativity in science.
Where are you originally from and what do you work on now?
I am from Canada. I am a theoretical biologist, with a focus on complex biological phenomena using evidence linked modelling. For my PhD thesis, I was trained in evolutionary biology and theoretical morphology, in the lab of Jon Stone at McMaster University, Canada. I developed a mathematical model that explores both growth and form of sea urchin skeletons. After graduating, I moved to Germany to work at the Max Planck Institute for Evolutionary Biology, where I was trained, under the supervision of Arne Traulsen, in evolutionary game theory. I developed models that capture and predict behaviours in complex social dilemmas such as climate change negotiations and host parasite interactions. Since 2016, I have been working as research associate, in the lab of Gary Bader, creating 3D mathematical models that explore cell development. My current research tries to understand how cell decisions happen and use mathematical modelling to predict how that affects cell fates during development.
Sea bloom, using wood burning
Were you always going to be a scientist?
No, my plan throughout high school was to become an engineer. I loved programming and I thought that engineering would be a good path, but then I discovered biology in my last term which led to a sudden change of heart and I switched to science!
Stem cells in bloom is a bouquet illustrating my first interpretation of the diversity stem cells in a developing system.
And what about art – have you always enjoyed it?
Yes, art has been a part of my life for as long as I can remember. My dad is an architect, so he sketched all the time and as soon I could hold a pencil, I sat next to him and sketched. During my teens, I learned to paint and that became my medium of choice. As an undergrad, I tried to take art classes in parallel to my bio degree, but all the classes conflicted with lab time so I couldn’t manage both and painting became a hobby.
Bouquet of bugs was a gift to an entomologist friend of mine, it is an acrylic painting and up close you will see it is not flowers but a collection of different insects
When did you started using science in your artwork?
As an undergrad, sketching my notes helped me retain information longer. Then, as a grad student, I helped develop an anatomy class that introduced students to sketching notes to learn and memorise information. Their incentive was that they were allowed to use their sketches during the infamous ‘bell-ringer’ exam. I now use my sci-sketchnotes as electronic summaries of research papers or talks.
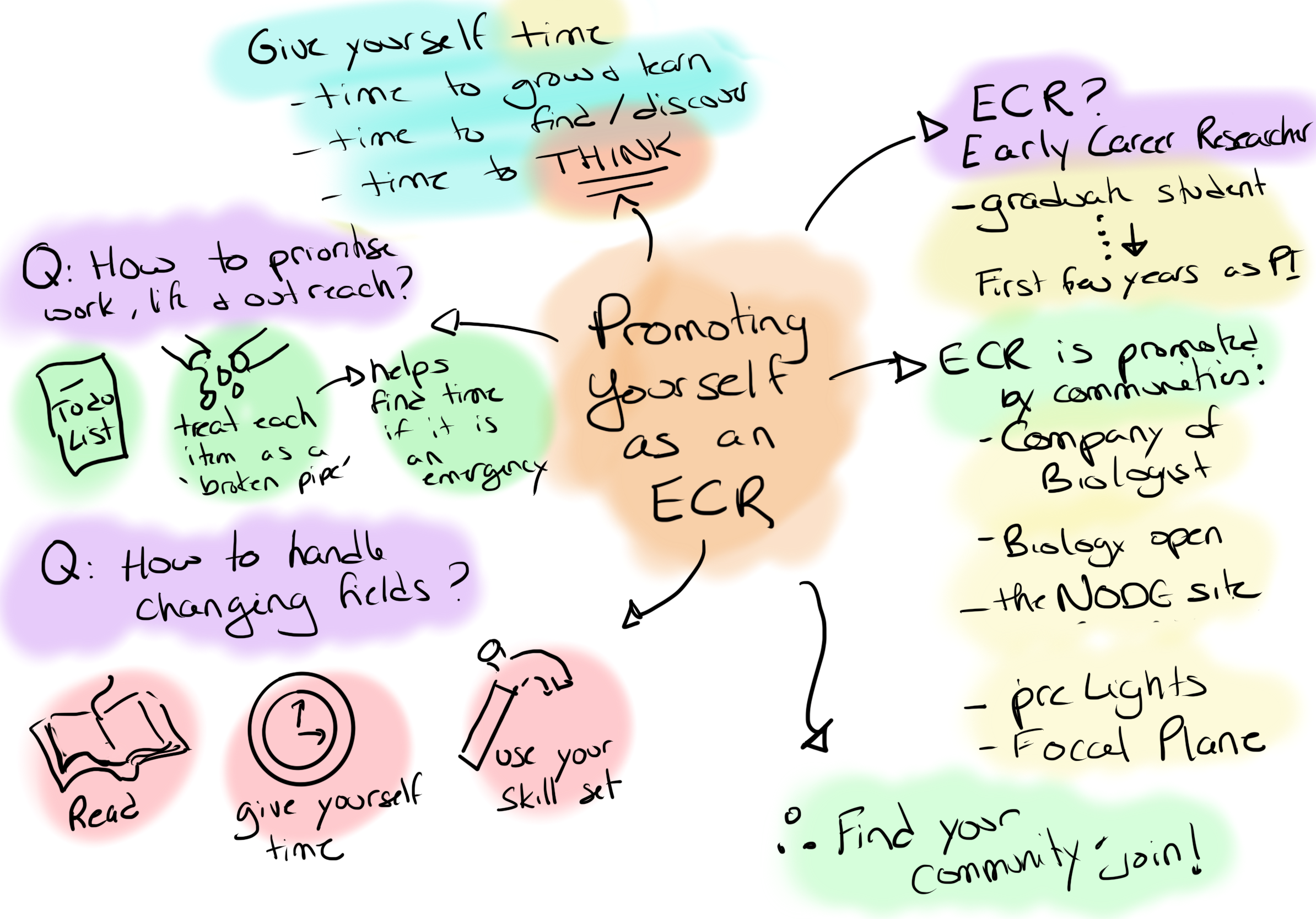
sci-sketchnotes from ECR advocacy event with The Company of Biologists
At the same time, it was hard to resist sketching the beautiful skeletons of the sea urchins that I studied during my PhD. My art then evolved into painting model organisms and other marine organisms. My supervisor and another professor appreciated them and asked to use them as the covers on the yearly reports for the department.
Acrylic painting for the biology department at McMaster (2005), I drew the model organisms (Arabidopsis, mouse, rat, frog, fruit fly, zebrafish and axolotl) trying to escape.
Does your art influence your science at all, or are they separate worlds?
I cannot imagine that it is possible to separate them. I think my creativity is one of the sources of my success in my research. I think having the ability to see things from a different perspective is an asset in science, and art helps me do that. A while back I read a piece by Sydney Brenner and a line stuck with me, ‘Science is the product of human minds, and the essence of research is creative innovation’ (Brenner, 1994). To me this quote says that you need both art and science, and that creativity is essential for discovery and progress in research.
SciArt poster that I created for a women in science meeting. For the central figure I chose Libby Hyman, a biologist who wrote the massive and beautiful echinoderm volumes which were an asset during my PhD.
You can find out more about Maria’s science and artwork on Twitter and on her webpage.
Thanks to Maria and all the other SciArtists we have featured so far.You can find the full list here. We’re always on the lookout for new people to feature in this series – whatever kind of art you do, from sculpture to embroidery to music to drawing, if you want to share it with the community just email thenode@biologists.com (nominations are also welcome!)
Registration is open for the 2022 Hydra Summer School in Stem Cell Biology and Regenerative Medicine.
This is a stimulating high-level course for postdocs, advanced PhD students (usually 2nd year & above), junior PIs and research-active clinicians. Places are limited and will be allocated on a competitive basis.
Leaders in the field will join you on the beautiful Greek island of Hydra to discuss topics ranging from recent advances in fundamental developmental and stem cell biology through to the latest breakthroughs in the clinic. There will also be sessions on ethics, research culture, careers, publishing, and outreach.
The Hydra summer school has been running since 2005 and has an outstanding reputation. Check out the testimonials from previous delegates: https://www.hydrasummerschool.org/testimonials
 (No Ratings Yet)
(No Ratings Yet)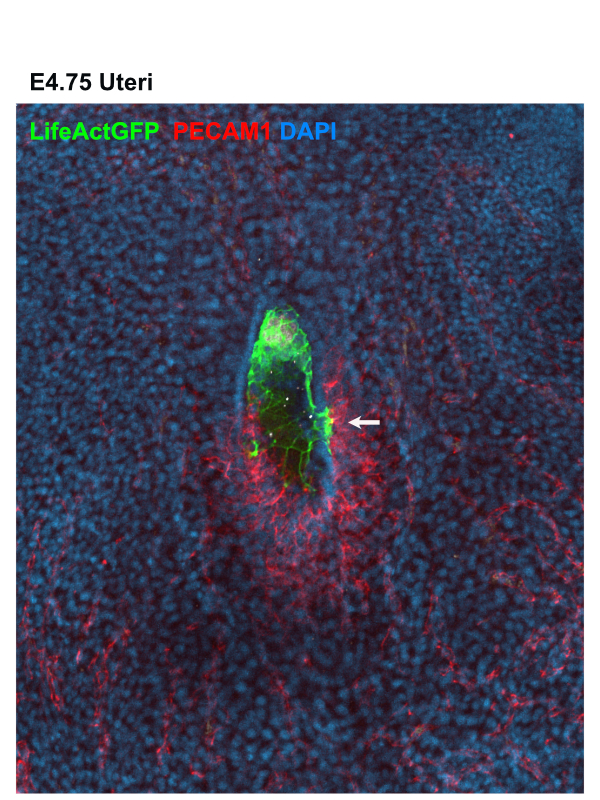
 (9 votes)
(9 votes)