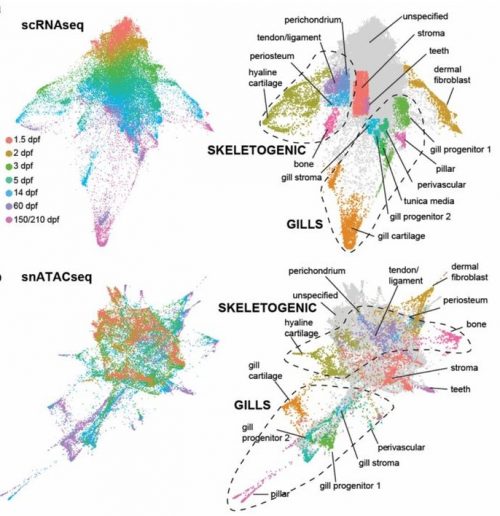
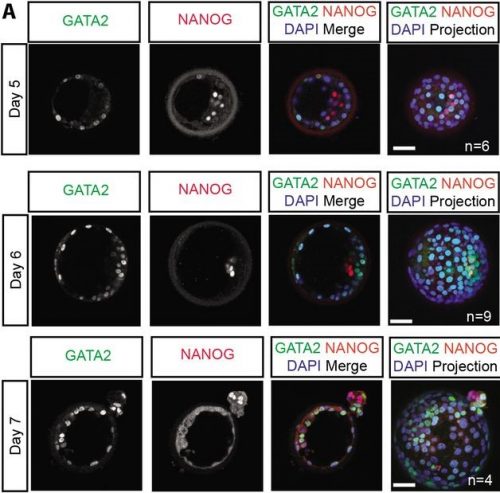
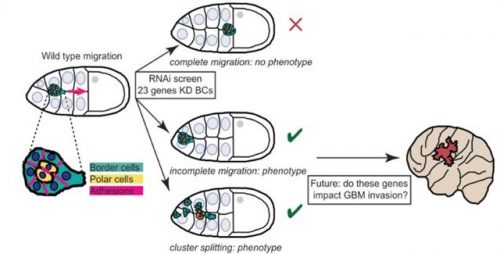
After the excitement and debate covered in the previous Developing news blog, the #devbio twittersphere was a little less controversial these past two weeks, but these are the topics we enjoyed.
Does collecting data drive biology forward?
An article in Nature discussing the importance of hypothesis driven science, not just data collection
Some thoughts from the #devbio community
The importance of theory and ideas (as well as data) for biology. By my @TheCrick colleague Paul Nurse
"Ideas, even tentative ones, are needed, along with the recognition that ideas will change as facts and arguments accumulate.”https://t.co/URohwnR6Ig
This x1000 – really excellent article. For example, there's currently too much single-cell-seq data in developmental biology without scientific ideas attached.
More papers with ideas in them! Not data for the sake of having data! Such an important and timely opinion article from Nobel Laureate Paul Nurse. I’m happy @nature published it and hope they’ll take it into consideration when evaluating submissions!!!https://t.co/fK0oN8Zgb4
Last week was postdoc appreciation week and we loved that our twitter feed was full of students, fellow postdocs and PIs giving thanks to their colleagues. We have collated all the tweets here
Careers tweets
We have also picked out three career-related tweets and the resulting discussions
Complain all you want, Twitter. I’m an academic and it’s the best job in the world. pic.twitter.com/CQogSyEuEl
Getting quite tired of the false choice narrative around Great Science vs Reasonable Pay in academia. Not helpful for trainees who are in their 20s/30s w loans and families. If you want to talk about diversity in STEM, start here
Thanks to the #DevBio community for sharing their thoughts, especially on twitter. If you have some news that you think we should share with the developmental biology community on our blog, please get in touch at thenode@biologists.com. If you are interested in getting involved with writing preLights you can find out more here.
On Wednesday 22 September Development hosted two researchers with interests in genes and development for our tenth Development presents… webinar
Below you’ll find each of the talks, plus a Q&A chaired by Development Editor Haruhiko Koseki. The next #DevPres webinar will be held on 13 October 2021, and chaired by James Briscoe – subscribe to our mailing list for updates.
Hirosuke Shiura (University of Yamanashi) – PEG10 viral aspartic protease domain is essential for the maintenance of fetal capillary structure in the mouse placenta
Thanks to everyone who responded to our call for shout out to, and stories about, the fabulous postdocs in your life. It was wondering to hear from students thanking their mentors, fellow postdocs appreciating the support and friendship of their colleagues and PIs praising their postdocs.
A few highlights from these posts were:
Steve Royle coining the term ‘postdoctacular’, which should hopefully gain traction in the science community
Andrew Bowman appreciating that his postdoc, Alonso, brings huge bottles of wine from Spain
Miguel Branco and Ondine Cleaver bringing our attention to all the different roles a postdoc has to fill
New PIs Maxim Greenberg and Gautam Dey recognising and appreciating the leap of faith a postdoc takes going to a new group (especially in the midst of a pandemic!)
We have included all the tweets we received below. They make this blog really long, but we love the sentiment behind them all, and wanted to give them a permanent place on the Node.
Of course, all these warm fuzzies are great, but interestingly our very first response was from Nishit Srivastava, raising the important subjects of contracts, living conditions and recognition for postdocs. A reminder that, as a community, to really show our appreciation to postdocs we must strive to support them all year around.
If institutes really want to appreciate postdocs, then walk the talk and show it in the form of better contracts, living conditions, and recognise us as the important cog in the wheel of science. https://t.co/SnfhmI8ccM
So appreciative and thankful for my two star postdocs Lomeli and Ezra! They make my science life better with their enthusiasm and ideas making it fun to come to the lab everyday. https://t.co/j8QeXNDczZ
I would like to thank Dr. @Marthabaydoun, my true mentor, my life savior, my inspiration and someone I learnt soooo much scientifically and personally. I hope the best, with all new opportunities and possibilities heading to her life. You go gurl 💪 https://t.co/Vtf2XMkA1a
as a young PI I was lucky to have @GiuGuiducci when I started my lab! She is not only doing amazing science but Is also a great colleagues and a mentor to other students + my right arm in the lab! Go girl! #PostdocAppreciationWeekhttps://t.co/bLWvpt1WxZ
#PostdocAppreciationWeek thanks to ll the postdocs who have inspired me since my undergrad to continue in research. Special thanks to one postdoc who throughout all the lockdowns supported my chemist without a lab phase, despite not being a chemist. 🤩 https://t.co/fxP6mZaeZI
Do not know where I would be without the postdocs I have worked w in my young career. My first two mentors, both postdocs, were exceptional humans and scientists. And in graduate school I’ve had @ZhaoyangLiu_5 and @elle_roberson to look up to as inspiring female scientists. https://t.co/VD2p9aKM2A
Grateful for my undergraduate postdoc mentor and stellar scientist @RobertJHuebner1 ❤️ and for @elle_roberson, the scientist who has inspired me with her work and female empowerment since the day we met ❤️ Thank you! https://t.co/l1DTU6jE5V
Postdoc life is not 'just' being at the bench focusing on your own experiments. The tireless @neverlethetruth and @smamante1 are also mentors, advisors, lab managers, administrators,… And somehow they keep coming to work with a smile! You guys rock. https://t.co/1ggnRbfg18
I don't know how I lucked into them, but @AnaMonteagudo4 and @jrichardalbert both went to a foreign country in the middle of a pandemic to work in a start-up lab. Not only are they great people, but now they are cranking out data! The appreciation (and pride) is immense! https://t.co/vYmTpZhwSR
Postdocs are leaders in the lab. They drive discovery, they mentor, and they think and work so darned hard. I have appreciated @XiaowuGu and @AhujaNH every single day. 🙏 The lab wouldn't be the same without them. https://t.co/Cvmc6ItHSq
In the latest Genetics Unzipped podcast, Dr Kat Arney takes a look at how we learned to read the book of life, from the earliest days of DNA sequencing to the very latest futuristic technologies.
Today, sequencing machines in labs around the world are churning out billions of basepairs of DNA data, and the very latest techniques even promise to allow us to read DNA inside cells. The cost of reading a whole human genome has gone from $2.7 billion dollars for the first one to a few hundred or even less or even zero, depending on how you count it and who’s paying.
And as you’ll know if you’re a fan of this podcast, widespread, cheaper access to rapid DNA sequencing has revolutionised almost every aspect of life sciences research, from personalized medicine and clinical diagnostics to conservation, infectious diseases, evolutionary biology and much, much more.
We’ve come a long way since those early days. So how did we get from there to here?
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
Royal Society Publishing has recently published a special issue of Philosophical Transactions B: Synchrony and rhythm interaction: from the brain to behavioural ecology compiled and edited by Michael D Greenfield, Henkjan Honing, Sonja Kotz and Andrea Ravignani and the articles can be accessed directly at www.bit.ly/PTB1835
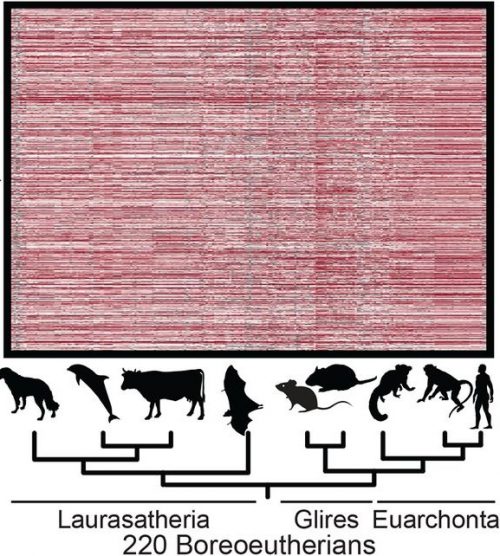
Which model is the best for studying human development?
A Perspective article in Nature about the Human Developmental Cell Atlas
Some responses from the #devbio community (note this article created a twitter storm, especially the size of the circles in Fig1c. We have picked out a few twitter threads)
The incredulous responses last week to chick embryos developing similarly to human at early stages explains A LOT. The reality is that all research models have benefits and drawbacks. There isn’t a “best one,” and if we don’t open our minds we miss out on cool discoveries. https://t.co/KsW9SA9TR5
While I do understand all the upset about the recent @Nature paper on the comparison of model oganisms, (see Figure 1c here : https://t.co/GI6WxmEe6i) I am not sure I would be upset for the same reason. Obviously fly is not the best system to reproduce human development, BUT…
There are many wonderful twitter threads offering advice to new graduate students, and indeed much of the advice is relevant to the wider community. Here are a couple of our favourites:
With the new academic year starting, and many people beginning fresh as graduate students, post-docs, or junior faculty, I have been thinking a lot about the core philosophies that govern my own perspectives on science. So I thought I would share. Welcome to The Tao of Cheese
Thanks to the #DevBio community for making this such an interesting couple of weeks, especially on twitter. If you have some news that you think we should share with the developmental biology community on our blog, please get in touch at thenode@biologists.com. If you are interested in getting involved with writing preLights you can find out more here.
For our tenth webinar in the Development presents… series, Development Editor Haruhiko Koseki (RIKEN Center for Integrative Medical Sciences) has invited interesting talks on genes and development.
Pablo Narravo Gil (Principal investigator at the Pasteur Institute) ‘H3K9me3 at Nanog, preparing and promoting the loss of pluripotency’
Hirosuke Shiura (Assistant Professor at the University of Yamanashi) ‘PEG10 viral aspartic protease domain is essential for the maintenance of fetal capillary structure in the mouse placenta’
The webinar will be held in Remo, our browser-based conferencing platform. After the talks you’ll have the chance to meet the speakers and other participants at virtual conference tables. If you can’t make it on the day, talks will be available to watch after the event on the Node. You can also sign up to our mailing list for email alerts.
Formerly known as Mas a Tierra and renamed in 1966, this small mountainous island is a remote tropical paradise known mostly for the fact that it’s said to be the inspiration behind Daniel Defoe’s novel Robinson Crusoe, hence the name.
Today, Robinson Crusoe Island is far-flung holiday destination for intrepid travellers, populated by a small island community of around 600. One day, about fifteen years ago, a woman named Pia Villanueva came to the island on holiday for a relaxing break – or, at least, that is until the islanders discovered that she was a speech therapist…
To tell the story, Kat’s joined by Dr Dianne Newbury, senior lecturer and principal investigator in the molecular genetics of speech and language at Oxford Brookes University.
She and her team have been working with the island population, which contains an unusually high number of people with speech and language impairment, to discover what their genes can teach us about speech and language development, and what happens when this goes awry.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
Welcome to our monthly scamper through the preprints on developmental biology and stem cells (and other related topics). Bumper section this month is the Genes & genomes section
The preprints this month are hosted on bioRxiv, arXiv and preprints.org – use these links to get to the section you want.
Phase transition specified by a binary code patterns the vertebrate eye cup Revathi Balasubramanian, Xuanyu Min, Peter M.J. Quinn, Quentin Lo Giudice, Chenqi Tao, Karina Polanco, Neoklis Makrides, John Peregrin, Michael Bouaziz, Yingyu Mao, Qian Wang, Bruna L Costa, Diego Buenaventura, Fen Wang, Liang Ma, Stephen H Tsang, Pierre J. Fabre, Xin Zhang
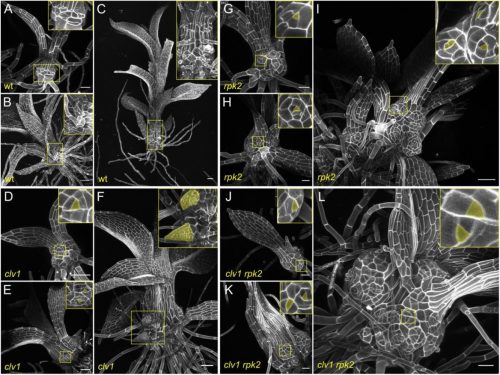
A gene regulatory network for neural induction Katherine E. Trevers, Hui-Chun Lu, Youwen Yang, Alexandre Thiery, Anna C. Strobl, Božena Pálinkášová, Nidia M. M. de Oliveira, Irene M. de Almeida, Mohsin A. F. Khan, Natalia Moncaut, Nicholas M. Luscombe, Leslie Dale, Andrea Streit, Claudio D. Stern
Integrated annotation and analysis of genomic features reveal new types of functional elements and large-scale epigenetic phenomena in the developing zebrafish Damir Baranasic, Matthias Hörtenhuber, Piotr Balwierz, Tobias Zehnder, Abdul Kadir Mukarram, Chirag Nepal, Csilla Varnai, Yavor Hadzhiev, Ada Jimenez-Gonzalez, Nan Li, Joseph Wragg, Fabio D’Orazio, Noelia Díaz, Benjamín Hernández-Rodríguez, Zelin Chen, Marcus Stoiber, Michaël Dong, Irene Stevens, Samuel E. Ross, Anne Eagle, Ryan Martin, Pelumi Obasaju, Sepand Rastegar, Alison C. McGarvey, Wolfgang Kopp, Emily Chambers, Dennis Wang, Hyejeong R. Kim, Rafael D. Acemel, Silvia Naranjo, Maciej Lapinski, Vanessa Chong, Sinnakaruppan Mathavan, Bernard Peers, Tatjana Sauka-Spengler, Martin Vingron, Piero Carninci, Uwe Ohler, Scott Allen Lacadie, Shawn Burgess, Cecilia Winata, Freek van Eeden, Juan M. Vaquerizas, José Luis Gómez-Skarmeta, Daria Onichtchouk, Ben James Brown, Ozren Bogdanovic, Monte Westerfield, Fiona C. Wardle, Carsten O. Daub, Boris Lenhard, Ferenc Müller
Somatic mutation rates scale with lifespan across mammals Alex Cagan, Adrian Baez-Ortega, Natalia Brzozowska, Federico Abascal, Tim H. H. Coorens, Mathijs A. Sanders, Andrew R. J. Lawson, Luke M. R. Harvey, Shriram G. Bhosle, David Jones, Raul E. Alcantara, Timothy M. Butler, Yvette Hooks, Kirsty Roberts, Elizabeth Anderson, Edmund Flach, Simon Spiro, Inez Januszczak, Ethan Wrigglesworth, Matthew W. Perkins, Robert Deaville, Megan Druce, Ruzhica Bogeska, Michael D. Milsom, Björn Neumann, Frank Gorman, Fernando Constantino-Casas, Laura Peachey, Diana Bochynska, Ewan St. John Smith, Moritz Gerstung, Peter J. Campbell, Elizabeth P. Murchison, Michael R. Stratton, Iñigo Martincorena
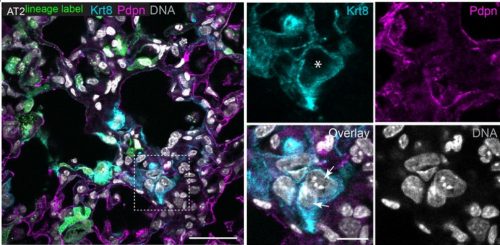
Kaplow, et al. used a CNN to predict brain OCR ortholog open chromatin status
A 3D transcriptomics atlas of the mouse olfactory mucosa Mayra L. Ruiz Tejada Segura, Eman Abou Moussa, Elisa Garabello, Thiago S. Nakahara, Melanie Makhlouf, Lisa S. Mathew, Filippo Valle, Susie S.Y. Huang, Joel D. Mainland, Michele Caselle, Matteo Osella, Stephan Lorenz, Johannes Reisert, Darren W. Logan, Bettina Malnic, Antonio Scialdone, Luis R. Saraiva
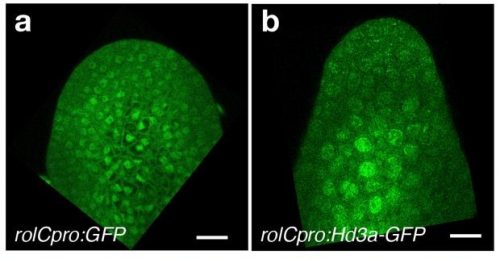
The control of transcriptional memory by stable mitotic bookmarking Maelle Bellec, Jeremy Dufourt, George Hunt, Helene Lenden-Hasse, Antonio Trullo, Amal Zine El Aabidine, Marie Lamarque, Marissa M Gaskill, Heloise Faure-Gautron, Mattias Mannervik, Melissa Harrison, Jean-Christophe Andrau, Cyril Favard, Ovidiu Radulescu, Mounia Lagha
Hematopoiesis at single cell resolution spanning human development and maturation Hojun Li, Jideofor Ezike, Anton Afanassiev, Laura Greenstreet, Stephen Y Zhang, Jennifer Whangbo, Vincent Butty, Enrico Moiso, Guinevere Connelly, Vivian Morris, Dahai Wang, George Q. Daley, Salil Garg, Stella Chou, Aviv Regev, Edroaldo Lummertz da Rocha, Geoffrey Schiebinger, Grant Rowe
Direct reprogramming of human embryonic to trophoblast stem cells Norah M.E. Fogarty, Ahmed Abdelbaki, Afshan McCarthy, Liani Devito, Alice E. Chen, Prabhakaran Munusamy, Paul Blakeley, Kay Elder, Phil Snell, Leila Christie, Paul Serhal, Rabi A. Odia, Mahesh Sangrithi, Kathy K. Niakan
Mesoderm-Derived PDGFRA+ Cells Regulate the Emergence of Hematopoietic Stem Cells in the Dorsal Aorta Vashe Chandrakanthan, Prunella Rorimpandey, Fabio Zanini, Diego Chacon, Young Chan Kang, Kathy Knezevic, Yizhou Huang, Qiao Qiao, Rema A. Oliver, Ashwin Unnikrishnan, Daniel R. Carter, Brendan Lee, Chris Brownlee, Carl Power, Simon Mendez-Ferrer, Grigori Enikolopov, William Walsh, Berthold Göttgens, Samir Taoudi, Dominik Beck, John E. Pimanda
Metabolic control of adult neural stem cell self-renewal by the mitochondrial protease YME1L Gulzar A. Wani, Hans-Georg Sprenger, Kristiano Ndoci, Srikanth Chandragiri, Richard James Acton, Désirée Schatton, Sandra M.V. Kochan, Vignesh Sakthivelu, Milica Jevtic, Jens M. Seeger, Stefan Müller, Patrick Giavalisco, Elena I. Rugarli, Elisa Motori, Thomas Langer, Matteo Bergami
Clonal dynamics of haematopoiesis across the human lifespan Emily Mitchell, Michael Spencer Chapman, Nicholas Williams, Kevin Dawson, Nicole Mende, Emily F Calderbank, Hyunchul Jung, Thomas Mitchell, Tim Coorens, David Spencer, Heather Machado, Henry Lee-Six, Megan Davies, Daniel Hayler, Margarete Fabre, Krishnaa Mahbubani, Fede Abascal, Alex Cagan, George Vassiliou, Joanna Baxter, Inigo Martincorena, Michael R Stratton, David Kent, Krishna Chatterjee, Kourosh Saeb Parsy, Anthony R Green, Jyoti Nangalia, Elisa Laurenti, Peter J Campbell
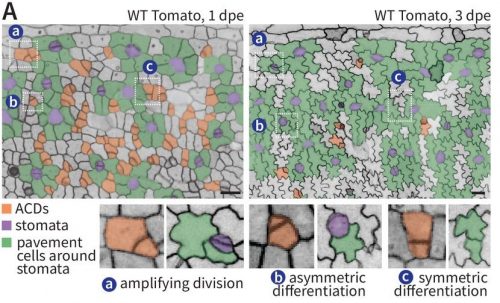
Image analysis of an Arabidopisis thaliana ovule using MorphographX 2.0 from Strauss, et al.
MorphoGraphX 2.0: Providing context for biological image analysis with positional information Soeren Strauss, Adam Runions, Brendan Lane, Dennis Eschweiler, Namrata Bajpai, Nicola Trozzi, Anne-Lise Routier-Kierzkowska, Saiko Yoshida, Sylvia Rodrigues da Silveira, Athul Vijayan, Rachele Tofanelli, Mateusz Majda, Emillie Echevin, Constance Le Gloanec, Hana Bertrand-Rakusova, Milad Adibi, Kay Schneitz, George Bassel, Daniel Kierzkowski, Johannes Stegmaier, Miltos Tsiantis, Richard S. Smith
Direct reprogramming of human fibroblasts into insulin-producing cells by transcription factors Marta Fontcuberta-PiSunyer, Ainhoa García-Alamán, Èlia Prades, Noèlia Téllez, Hugo Figueiredo, Rebeca Fernandez-Ruiz, Sara Cervantes, Carlos Enrich, Laura Clua, Javier Ramón-Azcón, Christophe Broca, Anne Wojtusciszyn, Anna Novials, Nuria Montserrat, Josep Vidal, Ramon Gomis, Rosa Gasa
Village in a dish: a model system for population-scale hiPSC studies Drew R. Neavin, Angela M. Steinmann, Han Sheng Chiu, Maciej S. Daniszewski, Cátia Moutinho, Chia-Ling Chan, Mubarika Tyebally, Vikkitharan Gnanasambandapillai, Chuan E. Lam, Uyen Nguyen, Damián Hernández, Grace E. Lidgerwood, Alex W. Hewitt, Alice Pébay, Nathan J. Palpant, Joseph E. Powell
Anatomical Structures, Cell Types, and Biomarkers Tables Plus 3D Reference Organs in Support of a Human Reference Atlas Katy Börner, Sarah A. Teichmann, Ellen M. Quardokus, James Gee, Kristen Browne, David Osumi-Sutherland, Bruce W. Herr II, Andreas Bueckle, Hrishikesh Paul, Muzlifah A. Haniffa, Laura Jardine, Amy Bernard, Song-Lin Ding, Jeremy A. Miller, Shin Lin, Marc Halushka, Avinash Boppana, Teri A. Longacre, John Hickey, Yiing Lin, M. Todd Valerius, Yongqun He, Gloria Pryhuber, Xin Sun, Marda Jorgensen, Andrea J. Radtke, Clive Wasserfall, Fiona Ginty, Jonhan Ho, Joel Sunshine, Rebecca T. Beuschel, Maigan Brusko, Sujin Lee, Rajeev Malhotra, Sanjay Jain, Griffin Weber
A Library of Induced Pluripotent Stem Cells from Clinically Well-Characterized, Diverse Healthy Human Individuals Christoph Schaniel, Priyanka Dhanan, Bin Hu, Yuguang Xiong, Teeya Raghunandan, David M. Gonzalez, Rafael Dariolli, Sunita L. D’Souza, Arjun S. Yadaw, Jens Hansen, Gomathi Jayaraman, Bino Mathew, Moara Machado, Seth I. Berger, Joseph Tripodi, Vesna Najfeld, Jalaj Garg, Marc Miller, Colleen S. Lynch, Katherine C. Michelis, Neelima C. Tangirala, Himali Weerahandi, David C. Thomas, Kristin G. Beaumont, Robert Sebra, Milind Mahajan, Eric Schadt, Dusica Vidovic, Stephan C. Schürer, Joseph Goldfarb, Evren U. Azeloglu, Marc R. Birtwistle, Eric A. Sobie, Jason C. Kovacic, Nicole C. Dubois, Ravi Iyengar
Mutationathon: towards standardization in estimates of pedigree-based germline mutation rates Lucie A. Bergeron, Søren Besenbacher, Tychele N. Turner, Cyril J. Versoza, Richard Wang, Alivia Lee Price, Ellie Armstrong, Meritxell Riera, Jedidiah Carlson, Hwei-yen Chen, Matthew W. Hahn, Kelley Harris, April Snøfrid Lo Natalie M Kleppe, Elora H. López-Nandam, Priya Moorjani, Susanne P. Pfeifer, George P. Tiley, Anne D. Yoder, Guojie Zhang, Mikkel H. Schierup
MITI Minimum Information guidelines for highly multiplexed tissue images Denis Schapiro, Clarence Yapp, Artem Sokolov, Sheila M. Reynolds, Yu-An Chen, Damir Sudar, Yubin Xie, Jeremy Muhlich, Raquel Arias-Camison, Milen Nikolov, Madison Tyler, Jia-Ren Lin, Erik A. Burlingame, Sarah Arena, Human Tumor Atlas Network, Young H. Chang, Samouil L Farhi, Vésteinn Thorsson, Nithya Venkatamohan, Julia L. Drewes, Dana Pe’er, David A. Gutman, Markus D. Herrmann, Nils Gehlenborg, Peter Bankhead, Joseph T. Roland, John M. Herndon, Michael P. Snyder, Michael Angelo, Garry Nolan, Jason Swedlow, Nikolaus Schultz, Daniel T. Merrick, Sarah A. Mazzilli, Ethan Cerami, Scott J. Rodig, Sandro Santagata, Peter K. Sorger
In the tenth SciArt profile of the series, we meet Elsa M. Quicazán-Rubio, a science communicator striving to bring the topic of biomimicry to a wider audience
Elsa participating in the Local Plastic Arts Fair (FLAP in Spanish) in Bogotá, 2021 as part of the creative collective Taller Casa Quemada (Photo by Camilo Buitrago)
Where are you originally from, where do you work now, and what do you work on?
I am from Colombia, where I studied my undergrad in Biology and wrote a thesis on biomechanics of hummingbird flight. This subject captivated me, and I completed a Masters degree at the University of California Riverside, USA, and an internship at the Wageningen University and Research, in The Netherlands. I then went back to The Netherlands to do a PhD in fish swimming.
During the pandemic, I was able to give shape to one of my dreams, Bioinspirada, a combination of Art and Science, connected mainly by Biomechanics and Biomimicry. I learnt how to make a website and launched Bioinspirada.com in January 2021. Since then, the ride has been awesome. I currently teach Biomimicry at the Colombian University EAN, collaborate in a team to develop Biomimicry workshops for kids at the Andes University and joined the creative space Taller Casa Quemada. I also participate as a speaker on subjects such as the role of females in science, biomimicry, and science communication.
Who hovers in the air? Hummingbirds. And who hovers in the water? Seahorses. What would the metamorphosis between a hummingbird and a seahorse look like? I was inspired by the work I did years ago on hummingbird flying, and the work on seahorse swimming during my PhD. I did this illustration in ink and colored pencils, using only three colors, 2017 – 2018
Has science always been an important part of your life?
Yes, science has always been an important part of my life. I remember talking to a friend in elementary school when we were about 10 years old, about our dream of becoming scientists. We both became scientists, one in medicine and the other in biology. I wanted to understand how animals work. Looking back at my childhood and youth, I see that the visits to my grandparents farm, and to the university, because of my parents’ jobs, contributed to this curiosity and help me to understand where I could get some of the answers.
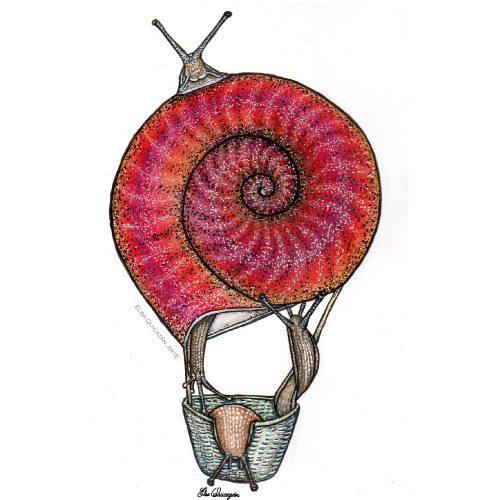
As I thought of snails, the question came to mind as to what other purposes could their shell have. What if they flew and the shell was like a balloon? This illustration turned out to be what they call in the cooking recipes a “deconstruction” of a snail in its parts Inks, colored pencils and watercolors, 2020
And what about art – have you always enjoyed being creative?
Since I was little, I have enjoyed painting and creating objects. In one of the kindergarten reports, the teacher refers to my ability to express myself better with drawings than words at the time. I read this just a couple of years ago, and it warmed my heart knowing that drawing has been there even before I can remember. During life I have had the support and guidance from different people. My parents encouraged me and supported me with courses and with their own creative inputs. I took art courses in and out of the University while I studied Biology.
Trees are more active than we generally think. They can communicate with each other. Now, on second thoughts, I think this illustration was inspired by the book “The Secret Life of Trees” by Peter Wohlleben. Color pencils and inks, 2020
What or who are your artistic influences?
Some of my artistic influences are Quino, a Latin American cartoonist with a very clean line. Rien Poortvliet, Alan Lee, and Brian Froud who illustrate natural and fantasy creatures. I especially remember a tales collection called “Cuenta Cuentos” (Salvat ed.) where each story was accompanied by rich visuals and a narrator’s recording. My classes with the Illustrator Esperanza Vallejo and Arts Profesor David Izquierdo had a special influence on my art because they both encouraged me and taught me how to explore and combine techniques, freeing my style.
After a while of moving between countries, cities and houses, I felt the urge to draw my roots. I could see that they travel with me and that they are also of many colors, they are happy, they are deep, and they are even a little strange. Ink and Ecolins (similar to watercolors), 2009 – 2010
How do you make your art?
Usually, I have the idea of illustrating an animal or a feeling and I just let that idea simmer in my subconscious for a while, until a shape comes up in my mind and I draft it. I often look at pictures of the animal that I want to represent and use them as references. Other times, I just have a quick idea and go drafting and finish the drawing over the draft itself. One of the illustrations that represent a feeling is the one with roots. This arose from the need to represent and see my own roots because at that time I was often moving from one country to another.
A fish that can fly using bird wings. I was inspired by the idea of teaching children about flying and swimming in animals. Color pencils and ink, 2019
Does your art influence your science at all, or are they separate worlds?
Up until recently my art and science were mostly separate, although I drew the biomechanics setups, which made them easier to build. But I feel that the freedom that I had while creating the figures of my first first-author paper on fish swimming allowed me to invite my art to dance with my science. I enjoyed the process so much and learnt tips and tricks of what a figure needs to have to be easily scannable, to be enjoyed by people with different color perceptions, and to tell a story by itself. I feel that figures can benefit so much from art and this combination converts them into an invitation for the public to approach our research.
“I feel that figures can benefit so much from art and this combination converts them into an invitation for the public to approach our research.”
Today, I work with art and science creating catchy images that invite conversation about what makes nature a powerful teacher. And I’m starting to collaborate with artists and engineers to enhance the public’s curiosity towards nature.
The Kingfisher, a small bird that hunts fish by diving into the water, is one of the inspirations for one of the fastest trains in the world. And one of the species of this little bird lives in the wetlands of Bogotá, Colombia. That’s how close we are to inspiration. This is a semi-mechanical version of this bird. Watercolors and inks. 2020
What are you thinking of working on next?
Right now, I am developing a course on Biomimetic Illustration which is about learning to illustrate and portray nature’s solutions to human and environmental challenges. One example of this kind of illustration is seen in the Semi-mechanic Kingfisher that I illustrated for a blog post in my webpage Bioinspirada.com. The Kingfisher is the inspiration for the high-speed train redesign because of the shape of its beak and head that allows for a smoother passage between media with different densities. Therefore, in the illustration I represent this beautiful bird with a bionic head and beak.
I also plan to bring illustration to teaching biomechanics and biomimicry to children. Because one of my mid-term goals is to foster a curiosity for nature-inspired design from childhood. This is how I want to contribute to promoting the care and research of nature in future generations.
We’re looking for new people to feature in this series throughout the year – whatever kind of art you do, from sculpture to embroidery to music to drawing, if you want to share it with the community just email thenode@biologists.com (nominations are also welcome!).
 (No Ratings Yet)
(No Ratings Yet)