In this episode of Genetics Unzipped we ask, what would have happened if Darwin had read Mendel? And what if they’d been on Twitter? Professor Greg Radick, a historian of genetics at the University of Leeds, has a few ideas. Plus, something else that Darwin would have loved – an ambitious project to sequence the DNA of everything across the tree of life from the team at the Wellcome Sanger Institute.
If you enjoy the show, please do rate and review and spread the word. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com
We are inviting applications for a 3-year PhD position in the general area of Cell Biology and Cell Biophysics, to commence in October 2019. The student will be physically based within the Department of Physiology, Development and Neuroscience (PDN, https://www.pdn.cam.ac.uk) at the University of Cambridge, UK. The Department has significant cell biology, biophysics and imaging infrastructure.
The proposed research project aims to explore how cells control their mechanical properties during cell shape changes. Cellular shape changes are driven by mechanical forces generated primarily by intracellular cytoskeletal networks. The project will focus on a key cytoskeletal network, the actin cortex, and investigate how cortex organization and mechanics are controlled at the molecular level. The project is interdisciplinary and will involve the combined use of live cell imaging, super-resolution microscopy, advanced electron microscopy, as well as molecular cell biology and cell biophysics techniques.
The cell cortex is a thin network of actin filaments, myosin and associated proteins that underlies the plasma membrane. The physical properties of the cortex, and primarily its contractile tension, determine the shape of animal cells. Precisely controlled changes in cortical contractility are at the basis of most cell deformations, and deregulation of cortical tension is implicated in a number of cancers.
The overall aim of the project is to understand how contractile tension is generated from the microscopic organisation of the cortical network. We will develop an interdisciplinary research plan to study:
– the spatial organization of cortical actin filaments, using cryo-electron microscopy and super-resolution optical microscopy,
– the biophysical basis of cortical contractility.
We will investigate these questions in the context of cell division, where the cortex reorganizes to create a tension gradient that drives cell cleavage during cytokinesis.
Candidates should have, or expect to obtain, a first class or upper second class honours degree in subjects such as Molecular Cell Biology or Biophysics. Students with a Physics degree who wish to work at the interface of physics and biology are also strongly encouraged to apply.
Funding is available to UK and EEA candidates. This project is a part of a collaboration with a partner at the University of Zurich (Switzerland) and will necessitate some international travel.
Suitably qualified and enthusiastic candidates interested in performing research in a dynamic multidisciplinary scientific environment should apply online via the University’s Applicant Portal for a PhD in Physiology, Development and Neuroscience (BLPN22).
List the name of your prospective supervisor Professor Ewa Paluch and the project title in the Research section of the form. You will also be required to submit a research proposal (1-2 pages of A4). An application is only complete when all supporting documents, including the 2 academic references, are submitted. It is the applicants responsibility to ensure their referees submit their references before the closing date Monday 27 May 2019.
Informal enquiries can be addressed to Prof. Ewa Paluch (ekp25@cam.ac.uk).
Our laboratory of synthetic development at IMBA (Vienna) is looking for a postdoctoral colleague to decode the self-organising behaviours of early embryos. This position is part of a HFSP with Shantanu Singh (Broad Institute) and Kyogo Kawaguchi (Riken Institute), and requires regular and frequent exchanges with these labs.
Requirements. We are looking for a colleague with:
a strong interest and experience in quantitative high-content imaging including significant experience with microscopy, biological assay development, and some experience using image processing software on biological assays
an ability to clearly communicate with biology collaborators about project aims and results, and with software engineers about image processing and software needs
a personality that is helpful, professional, highly organized, and goal-oriented
useful but not required: stem cell culture experience, CellProfiler expertise, programming experience (e.g., Python), and experience with Linux environments, cloud computing clusters, and bioinformatics/statistics tools (e.g., R).
About us. Our laboratory gathers stem cell biologists, geneticists, and engineers who study organizing principles in the early embryo. We created the blastoid system, a currently unique in vitro model of the pre-implantation conceptus generated solely from stem cells [Nature 2018]. Upon transfer in utero, blastoids implant and initiate a pregnancy. We also develop microsystems, genetic and computational tools to modulate and analyse self-organisation. We are conscientious, enjoy a professional, no-fuss environment, and are passionate about what we do.
About IMBA. The IMBA is an institute of the Austrian Academy of Science and of the Vienna Biocenter. It hosts 15 research groups that foster strong creativity and world-class science in biology, including stem cells, organoids and development. IMBA and the Vienna Biocenter provide the utmost technical support to researchers, including in microscopy (e.g., lattice, mesoscale and macroscale LSFMs handled by technical experts), so that you can focus on the science. See https://www.imba.oeaw.ac.at/scientific-facilities/
We are seeking to recruit a new member to our team at the University of Cambridge that contributes to two Drosophila databases: Virtual Fly Brain (VFB; http://virtualflybrain.org) and FlyBase (http://flybase.org/). Virtual Fly Brain is a resource that helps Drosophila neuroscientists map and visualize neuronal circuits, and find the experimental reagents to manipulate the circuits to reveal their role in perception, learning, memory and complex behaviours. The successful applicant will aid this process in three ways: 1) by annotating the expression of Gal4 drivers within the adult brain, using data from large scale datasets and the primary research literature; 2) assisting in the integration of data from single-cell RNA sequencing datasets; and 3) leading the effort to assess the functionality of improvements to the website, by recruiting members of the local neuroscience community and working with them and other VFB members to perform usability testing of the website. Much of the curated data will also be integrated into FlyBase, the primary community resource for Drosophila genetic, genomic and functional data. You will interact day to day with the rest of the expert scientist-curators in the Virtual Fly Brain and FlyBase team, housed in the Department of Physiology, Development and Neuroscience, and VFB colleagues in the Cambridge area and Edinburgh. Members of VFB and FlyBase will provide appropriate training.
The Mokalled lab in the Department of Developmental Biology at Washington University School of Medicine is seeking highly motivated scientists to join our vibrant and collaborative research team (http://www.mokalledlab.com/). We use zebrafish as a discovery platform to study spinal cord regeneration across species. Candidates with expertise in zebrafish regeneration models or cell reprogramming models are encouraged to forward a cover letter, CV, and list of 2-3 references to mmokalled@wustl.edu.
The Young Tunisian Researchers in Biology Network:
Tunisia is suffering brain drain, especially post Tunisian-revolution (2011). Recent non-official reports say that over 90.000 senior quitted Tunisia since 2012, most of them are doctors, pharmacists and engineers. Aware of considerable role that may Tunisian scientist diaspora could play in the development of Tunisia in general and science and education in particular, the YTRB project was born (Young Tunisian Researchers in Biology). The project is all the more credible as all the founders were graduated from Tunisian universities and spent at least one post.doc outside Tunisia. The idea came out in early 2015 during a biology conference in Tunisia. Young Tunisians post.doc working in France, Tunisia, Sweden and Germany discussed the opportunity to organise a scientific event in Tunisia dedicated to young principal investigator (PI) as speakers essentially. The first YTRB committee was created with a fixed target: May 2016 will be the date of the 1st International Symposium of Young Researchers in Biology in Tunisia. We had an excellent speakers selection from Tunisia, Brazil, Europe and Asia. We had also a plenty of short oral talks and posters presented by local and international participants and definitely putting the event as the major scientific symposium in biology ever organised in Tunisia. Encouraged by this big success, we decided to continue building the Network. The major aim of YTRB is to create a new and young collaboration network and to help young Tunisian colleagues to connect to their peers all over the world, in order to provide knowledge, experience and technology exchange. The main activities of the YTRB Network are first conducting biannually an international biology symposium for young researchers. In collaboration with Tunisians universities and faculties, the organisation of several workshops in different thematic, Skype conferences and webinars, technology advances days wish are a mini symposium with focus on single thematic. Moreover, we created an annual science vulgarisation prize to help young Tunisian scientists to communicate their findings to a large public. In summary, we believe that YTRB is creating a new science climate in Tunisia marked by increasing the meetings, symposia, workshops and even chair in universities with the tag “young scientist” since the first symposium May 2016. This definitely encourages us to continue our work and investment to contribute modestly renewing the scientific research and build together the future science in Tunisia with the motto: Be Young, be Brilliant, Be YTRB!
Audience during the first international symposium of the YTRB Network, Monastir, Tunisia, May 2016
Organisers and volunteers during the second international symposium of the YTRB Network, Hammamet, Tunisia, May 2018
Our Thanks To
None of the work by the YTRB Network would be possible without the amazing devotion of Chamseddine Kifagi, Myriam Fezai, Fakher Frikha, Imen Chamkha, Walid Elfalleh, Radhouène Doggui, Ons Mamai, Chaker Slaymi, Radhouène Doggui, Nourhène Fendri, Abdelaziz Tlili, Alif Chebbi, Azza Sellami, Hana Trigui, Dhouha Daassi and many others. In addition, the YTRB activities could not be performed without the generous help from various organisations and scientists. I would like to thank the German, Italian, Spanish and French cultural institutes in Tunisia. Moreover, I’m grateful to the MNS” (Mediterranean Neuroscience Society, https://mnsociety.live/) and DrosAfrica (http://drosafrica.org/). We encourage every reader of this article to visit our website (https://ytrbiology.weebly.com/) and help the Network from their own work and time with any advertisement and financial support.
A postdoc position is available in the Laird lab at the University of California, San Francisco to study cell heterogeneity during development using quantitative imaging, lineage tracing and single cell approaches in mouse and ESC models. Our collaborative team focuses on the basis of heterogeneity in the developing germline and its consequences for gamete function. The position is in the UCSF Edythe and Eli Broad Center for Regeneration Medicine and Stem Cell Research located at the UCSF Parnassus Heights campus, in the heart of San Francisco. UCSF offers an outstanding developmental biology community, access to cutting edge technologies and a supportive working environment. Candidates with a Ph.D. degree in a biological science, publications, demonstrated creativity and research experience in a relevant field such as genetics, biochemistry, live imaging or basic bioinformatics should submit a C.V. and names of at least 2 references via email to diana.laird@ucsf.edu.(more…)
Fully funded postdoctoral positions are presently available in the Conlon Lab, whose studies focus on identifying the molecular networks that are essential for early heart development and how alterations in these networks lead to congenital heart disease and sexual dimorphism. For these studies, we use a highly integrated approach that incorporates developmental, genetic, proteomic, biochemical and molecular-based studies in mouse, Xenopus and stem cells.
Recent advances and projects of interest in the Conlon lab include studies that define the cellular and molecular events that lead to cardiac septation, those that explore cardiac interaction networks as determinants of transcriptional specificity, the mechanism and function of cardiac transcriptional repression networks, and the regulatory networks of cardiac sexual dimorphism.
Candidates should have recently obtained or be about to obtain a Ph.D. or M.D. in a field of biological science and should have a strong publication record. Outstanding and highly motivated candidates should apply by email to Dr. Frank L. Conlon (frank_conlon@med.unc.edu) and include a CV/resume, three references and description of your specific interest in our research programs.
In the mid-1900s, Conrad Hal Waddington introduced the idea of development as a series of branching decisions taken under the control of genes1. In mammals, the first of these decisions takes place before the implantation of the embryo in the maternal uterus and leads to the distinction between the trophectoderm (TE, future placenta) and the inner cell mass (ICM, future embryo and yolk sac). In our recent study, we dissected the role of the Notch signalling pathway during early preimplantation development and found that it promotes the gradual loss of potency prior to the first lineage choice.
This project started back in 2014, after our previous work led by Teresa Rayon in which we characterised a regulatory element upstream of Cdx2, key gene in the specification of the trophectoderm2. Cdx2 was known to act downstream of the Hippo pathway, but we found that this enhancer was activated by the convergence of two pathways: Hippo and Notch. I had recently started my PhD in Miguel Manzanares’ lab at the Centro Nacional de Investigaciones Cardiovasculares (CNIC) in Madrid, and we decided to explore in more detail what Notch was doing in these early stages of mouse development.
Different labs had been studying the role of the Hippo pathway during preimplantation development for a few years. So, we recapitulated all the information to try to find a cue of how Notch could be cooperating with it. Hippo is known to act as a readout of cell polarity and thus, only polarised cells (facing the outside) allow YAP to bind the transcription factor TEAD4 and activate Cdx2. However, initial expression of Cdx2 in the early compacted morula occurs in both inner and outer cells. Also, in the original work by Hiroshi Sasaki’s lab3, the authors claimed that the drop of Cdx2 expression in Tead4 mutant embryos was more dramatic in the blastocyst, when Cdx2 is only in the polarised TE, than in the morula. Therefore, our first hypothesis was that Notch could be acting in the early phases of Cdx2 expression, when this expression could not be completely explained by the role of YAP in polarised cells. We checked CDX2 in Rbpj and Notch1 mutant morulae (transcription factor and receptor of the Notch pathway respectively) and interestingly, we saw that CDX2 was strongly diminished specifically in the early morulae (<16 cells). After that, CDX2 levels were recovered, presumably because of the action of the Hippo pathway. Only double mutants for both Rbpj and Tead4 completely lacked CDX2 expression in the morula (and they did not reach the blastocyst stage). Cdx2 was responding to both pathways, but it seemed to behave differently depending on the stage. To be able to modulate the action of each pathway and verify if they were acting differently in these time windows, we used pharmacological inhibitors that allowed us to block them in a time-controlled manner. In the morula stage, only the blockade of the Notch pathway affected the expression of Cdx2. In contrast, the inhibition of TEAD/YAP was the one reducing Cdx2 from morula to blastocyst. Thus, we confirmed that although both pathways converged to regulate the same target gene, they did not act the same way.
CDX2 expression in wildtype and Rbpj mutant morulae. Reduction of CDX2 is evident in the early stages of Rbpj mutant embryos (left panel, 8-16 cells).
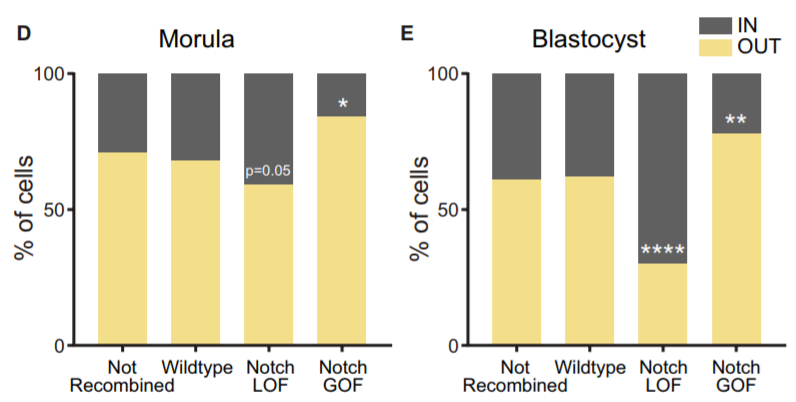
Another aspect we wanted to study was how Notch behaved during the morula to blastocyst transition, when its activity gets gradually restricted to the TE. In 2015, I was awarded a short stay fellowship from the Spanish Government that allowed me to visit the lab of Kat Hadjantonakis at the Sloan-Kettering Institute in New York for a few months, to continue with our collaboration and perform live imaging in embryos from the Notch activity reporter line (CBF1-VENUS)4. After some struggle, and with the help of Min Kang, we finally got movies where the embryos stayed in focus during the whole time-lapse (~24 hours). But the tougher part appeared later, back in Madrid. How could we manage all the information from those movies? The cell tracking for each embryo took a while… a long while. And once it was done, we still needed to consider many aspects. Fortunately, Antonio Lopez-Izquierdo, a biomedical engineer student, joined the lab to carry out the final project for his degree. He could programme in Matlab, so he developed a tool to 3D-reconstruct the embryos in each time frame and analyse the behaviour of the intensity of the reporter according to the position of the blastomeres within the embryo. The results indicated that there were already differences in the reporter intensity levels between outer and inner blastomeres in the morula, and that there was some correlation between these intensity levels and the position that cells occupied within the embryo which required further study. By then, the laboratory of Rui Benedito (our neighbours next door) had generated a transgenic line to produce mosaic cell populations with different Notch activity levels upon specific LoxP/CRE recombination5. We used this system to confront wildtype blastomeres with Notch loss of function (LOF) and gain of function (GOF) blastomeres. Beautifully, we saw how these differences affected the positioning of the cells within the embryo: Notch GOF blastomeres were more prone to occupy outer positions at the expenses of Notch LOF blastomeres which preferentially occupied inner locations.
Percentage of not recombined cells or recombined for each cassette from the iChr-Notch-Mosaic line (wildtype, Notch LOF or Notch GOF) that are in an inner or outer position at the morula and blastocyst stage.
At that point, we knew more details about the role of Notch regulating Cdx2 and favouring the positioning of blastomeres in the embryo. Nevertheless, we had the feeling that Notch was doing something else and we wished to gain a broader view on how it was working. Transcriptome profiling using low amounts of RNA was emerging, so that was the way to go. We decided to carry out single embryo RNA-seq in wildtype and Rbpj mutant morulae. Two issues arose: each sample consisted of only a dozen cells so we had to test and fine-tune the protocol to make sure that it would work with the limited material; and we could only genotype after the sequencing so we did not know how many samples of each genotype we had. Once we had it, the analysis showed that most of the genes (~70%) were downregulated; these were not only TE-related genes, but also some pluripotency genes. So, the role of Notch did not seem to be just the specification of the TE.
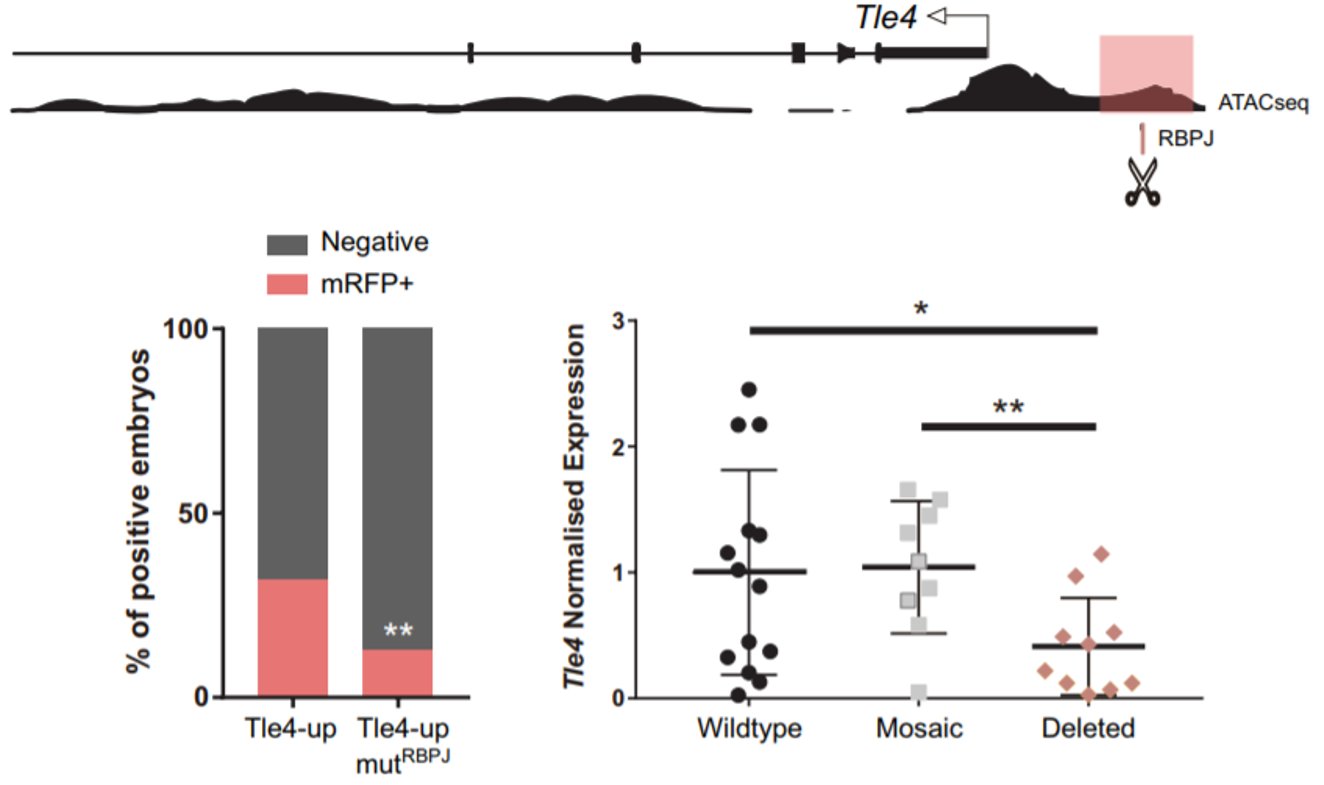
Remarkably, among the genes that were upregulated, we found two naïve pluripotency genes: Prdm14 and Dppa3. When we obtained this result, we had already seen that Notch was active very early, when the embryo consists of only four cells. Interestingly, Prdm14 is known to be heterogeneously expressed at the 4-cell stage and its expression fades away to be re-expressed in the ICM of the blastocyst6. That is the opposite pattern of Notch activity. Hence, we hypothesised that Notch could be blocking the expression of those naïve pluripotency genes in the early embryo, to boost the first differentiation programs. We did not think that this could be a direct target of Notch, given that when Notch is active it mainly activates gene expression. It is when Notch is not active that RBPJ can act as a repressor. We wondered if we could identify any gene that could be activated by Notch to repress the naïve pluripotency genes. To do that, we combined the analysis of our RNA-seq with predicted RBPJ binding motifs and data of ATAC-seq in 8-cell embryos7. In the resulting list (which was much more bearable), we found two candidates that had been described to block naïve pluripotency markers in mouse ES cells: Tle4 and Tbx3; and we decided to study them in parallel. With the help of Isabel Rollan in the lab, we identified two enhancers in the genomic landscapes of those genes that included an RBPJ motif in their sequences (Tle4-up and Tbx3-i7). To see if these sites were important for the regulation of the enhancers, we mutated these motifs and only the activity of the Tle4 enhancer was reduced. Finally, in order to see if these sites also affected the endogenous expression of Tle4 and Tbx3 respectively, we deleted them using the CRSPR/Cas9 system and we observed that Tle4 expression was affected in the edited embryos, but Tbx3 levels remained normal. Thus, RBPJ was important for the activity of the Tle4-up enhancer and for proper Tle4 expression in the morula.
An RBPJ binding motif upstream of Tle4 is important for proper activity of the enhancer in which is included (highlighted in pink). Deletion of this site by CRISPR/Cas9 decreases Tle4 endogenous expression (bottom right).
We concluded that Notch was regulating the transition in the embryo towards the first lineage decision. First, promoting a differentiating scenario by blocking naïve pluripotency genes (possibly through TLE4) and then, inducing the expression of Cdx2 to specify the trophectoderm in cooperation with Hippo.
It is curious to think how the role of Notch during preimplantation development was ruled out more than a decade ago because single mutants were not lethal until postimplantation stages, and how convergence with other inputs can give robustness to embryonic development albeit masking important roles.
I want to finish thanking other lab members who were also involved in this exciting story: Mariajo Andreu, Julio Sainz de Aja and Javier Adan.
Sergio Menchero, Isabel Rollan, Antonio Lopez-Izquierdo, Maria Jose Andreu, Julio Sainz de Aja, Minjung Kang, Javier Adan, Rui Benedito, Teresa Rayon, Anna-Katerina Hadjantonakis, Miguel Manzanares
eLife 2019;8:e42930 DOI: 10.7554/eLife.42930
References:
Slack, J. M. W. Conrad Hal Waddington: the last Renaissance biologist? Nat. Rev. Genet.3, 889–895 (2002).
Rayon, T. et al. Notch and hippo converge on Cdx2 to specify the trophectoderm lineage in the mouse blastocyst. Dev Cell30, 410–422 (2014).
Nishioka, N. et al. Tead4 is required for specification of trophectoderm in pre-implantation mouse embryos. Mech Dev125, 270–283 (2008).
Nowotschin, S., Xenopoulos, P., Schrode, N. & Hadjantonakis, A. K. A bright single-cell resolution live imaging reporter of Notch signaling in the mouse. BMC Dev Biol13, 15 (2013).
Pontes-Quero, S. et al. Dual ifgMosaic: A Versatile Method for Multispectral and Combinatorial Mosaic Gene-Function Analysis. Cell170, 800–814.e18 (2017).
Burton, A. et al. Single-Cell Profiling of Epigenetic Modifiers Identifies PRDM14 as an Inducer of Cell Fate in the Mammalian Embryo. Cell Rep.5, 687–701 (2013).
Wu, J. et al. The landscape of accessible chromatin in mammalian preimplantation embryos. Nature534, 652–657 (2016).
3-year fixed term postdoctoral position funded by the MRC
About the position:
The Percharde lab at the MRC London Institute of Medical Science (MRC LMS) is looking to recruit 1-2 talented and highly-motivated postdocs to join our group. The Chromatin & Development group is a recently-established team focused on understanding the molecular events surrounding cell fate choices during early development.
Our recent research indicates that Transposable Elements (TEs) have important, undiscovered roles during development, and projects in the lab include uncovering how TE regulation is important during embryogenesis, as well as how unrestrained TE expression may contribute to cases of disease. To answer these questions, we routinely use mouse and human embryonic stem cells (ESCs) and mouse embryos as models, adopting a combination of genome-wide techniques, CRISPR/Cas9 technology, high-resolution microscopy, biochemistry and molecular embryology approaches to probe gene and TE function. Visit www.perchardelab.com and https://lms.mrc.ac.uk/research-group/chromatin-and-development/
for more information. Closing date 16th May, interviews on 3-5th June 2019.
Candidate specifications:
Candidates should have a PhD or be in the final stages of completing one, and have a strong background in epigenetics, developmental biology and/or gene or transposon regulation. They should also have one or more first-author publications published or being submitted to a peer-reviewed, internationally recognised journal. Experience working with mouse and/or human ESCs/iPSCs or mouse embryos is highly desirable, as well as an interest in transposon biology.
About the institute:
This is an MRC Postdoctoral Scientist position providing training and development supporting post-doctoral scientists in early or changed career paths helping establish them as successful research scientists in their chosen field.
The MRC London Institute of Medical Sciences is a research institute within UK Research and Innovation. UK Research and Innovation is a new entity that brings together nine partners to create an independent organisation with a strong voice for research and innovation, more information can be found at www.ukri.org
Apply:
For full details of this post and to complete an online application, visit: https://mrc.tal.net/vx/lang-en-GB/appcentre-1/candidate/postings/1195 and upload your CV, the names and contacts of two scientific references, along with a cover letter stating why you are applying for this post (providing evidence against the requirements as per the Job Description and Person Specification). Please quote reference number LMS – 1071.
 In this episode of Genetics Unzipped we ask, what would have happened if Darwin had read Mendel? And what if they’d been on Twitter? Professor Greg Radick, a historian of genetics at the University of Leeds, has a few ideas. Plus, something else that Darwin would have loved – an ambitious project to sequence the DNA of everything across the tree of life from the team at the Wellcome Sanger Institute.
In this episode of Genetics Unzipped we ask, what would have happened if Darwin had read Mendel? And what if they’d been on Twitter? Professor Greg Radick, a historian of genetics at the University of Leeds, has a few ideas. Plus, something else that Darwin would have loved – an ambitious project to sequence the DNA of everything across the tree of life from the team at the Wellcome Sanger Institute. (No Ratings Yet)
(No Ratings Yet)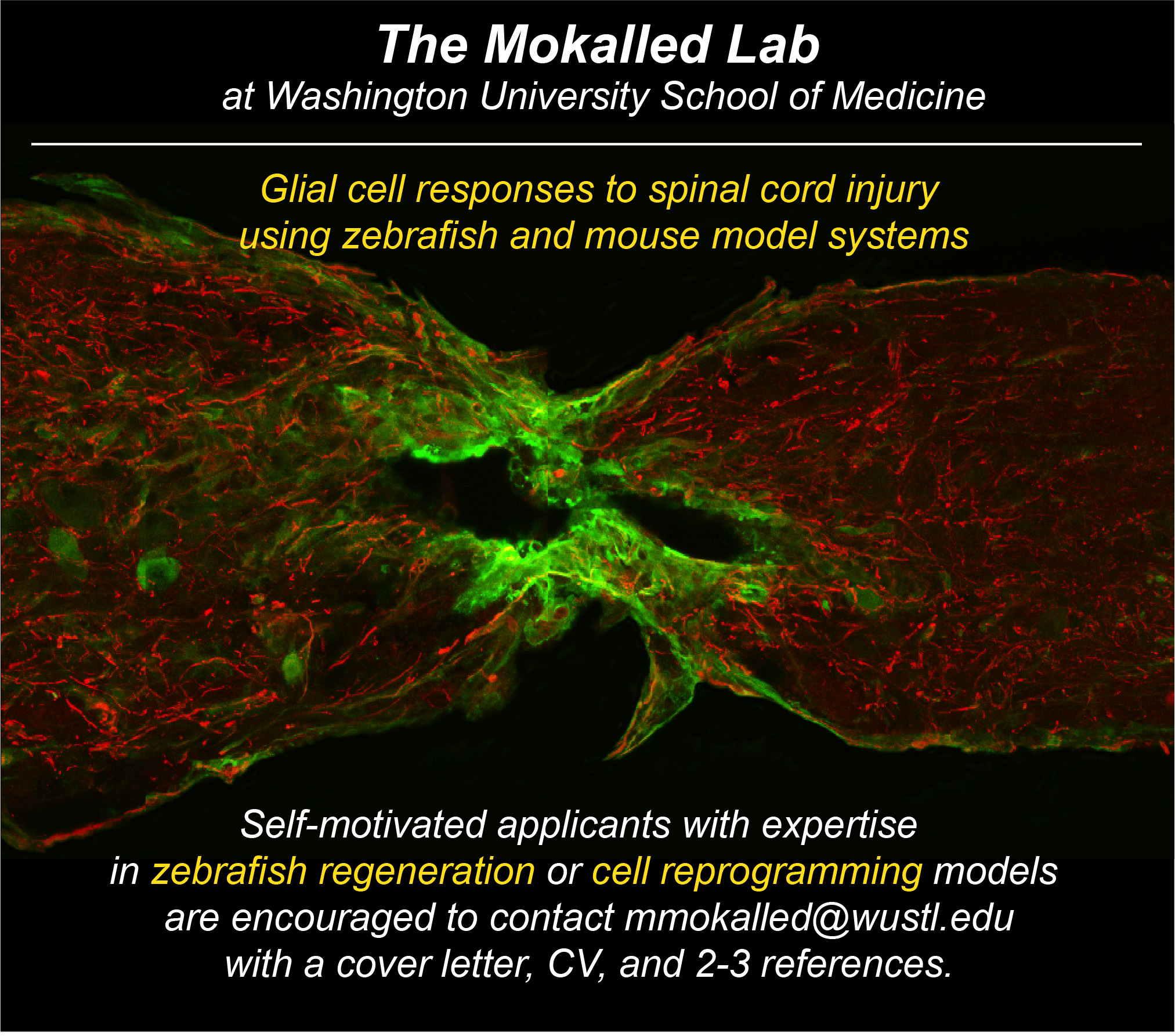


 (7 votes)
(7 votes) San Francisco to study cell heterogeneity during development using quantitative imaging, lineage tracing and single cell approaches in mouse and ESC models. Our collaborative team focuses on the basis of heterogeneity in the developing germline and its consequences for gamete function. The position is in the UCSF Edythe and Eli Broad Center for Regeneration Medicine and Stem Cell Research located at the UCSF Parnassus Heights campus, in the heart of San Francisco. UCSF offers an outstanding developmental biology community, access to cutting edge technologies and a supportive working environment. Candidates with a Ph.D. degree in a biological science, publications, demonstrated creativity and research experience in a relevant field such as genetics, biochemistry, live imaging or basic bioinformatics should submit a C.V. and names of at least 2 references via email to diana.laird@ucsf.edu.
San Francisco to study cell heterogeneity during development using quantitative imaging, lineage tracing and single cell approaches in mouse and ESC models. Our collaborative team focuses on the basis of heterogeneity in the developing germline and its consequences for gamete function. The position is in the UCSF Edythe and Eli Broad Center for Regeneration Medicine and Stem Cell Research located at the UCSF Parnassus Heights campus, in the heart of San Francisco. UCSF offers an outstanding developmental biology community, access to cutting edge technologies and a supportive working environment. Candidates with a Ph.D. degree in a biological science, publications, demonstrated creativity and research experience in a relevant field such as genetics, biochemistry, live imaging or basic bioinformatics should submit a C.V. and names of at least 2 references via email to diana.laird@ucsf.edu.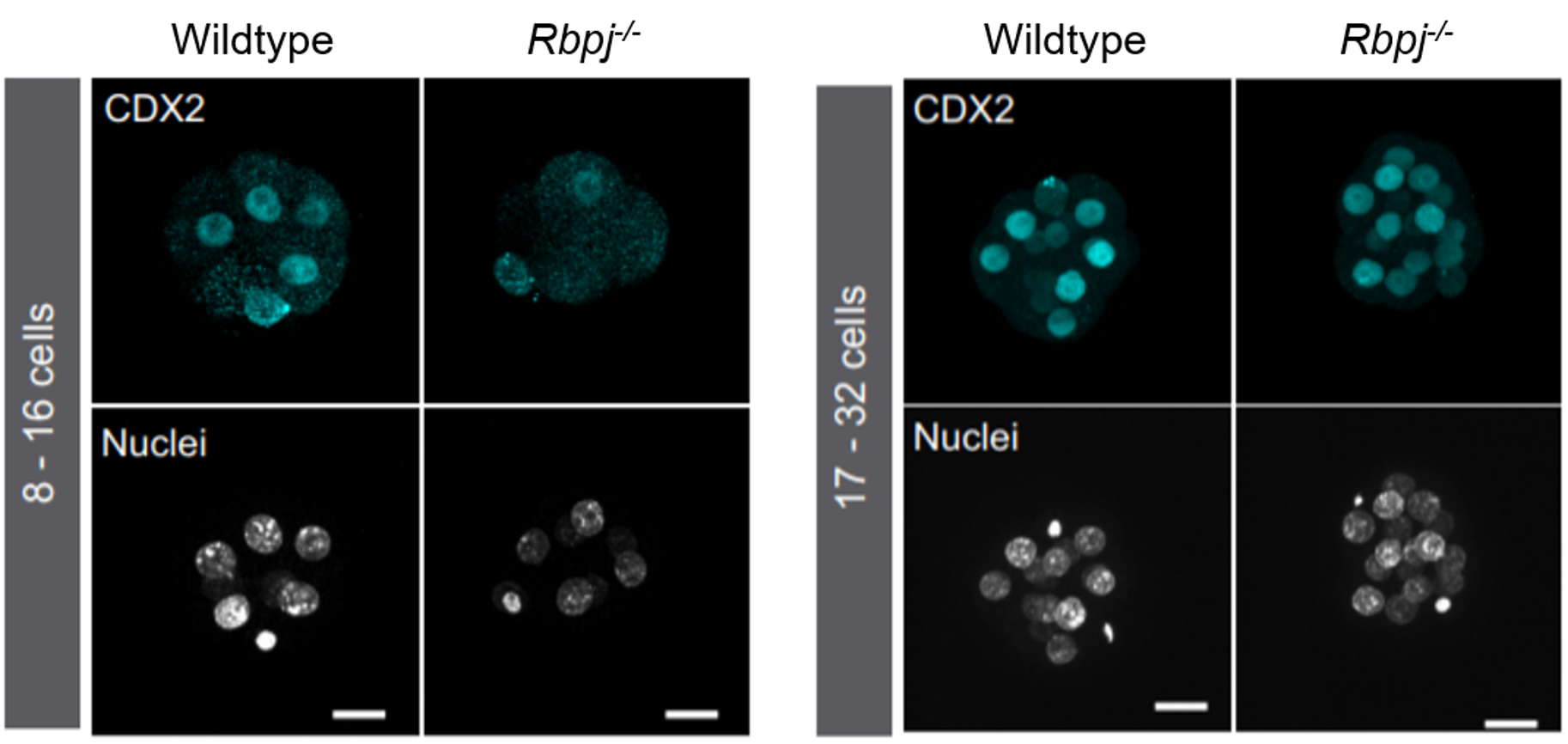
 (1 votes)
(1 votes) Our
Our