The Developmental Biology Unit seeks to understand the general principles and mechanisms underlying the development of multicellular organisms. Researchers in the unit combine the power of genetic model organisms with quantitative imaging and -omics technologies, synthetic biology, reduced (in vitro) systems and theoretical modelling, to create a cross-cutting approach to modern developmental biology.
Research in the Developmental Biology Unit is firmly embedded within the overall EMBL environment, with extensive in-house collaborations, access to outstanding graduate students and postdoctoral fellows, and support from cutting-edge facilities, including genomics, transgenesis, metabolomics, mass-spectrometry, and microscopy. EMBL brings together the most talented scientists, empowering them to explore bold new areas of biological inquiry and carry out interdisciplinary research.
We are seeking to recruit outstanding group leaders who aim to establish novel approaches to investigate multicellular development at all scales, from the cellular and tissue, to the whole organism level.
For more information and for the application please go to the following links:
Axon guidance relies on the reception and integration of molecular cues from the environment by growth cones, and defective pathfinding results in misplaced projection patterns in the mature nervous system. A new paper in Development investigates this process in the Drosophila neuromucular system, as well as the consequences of axonal miswiring to locomotion. We caught up with lead author Jaqueline Kinold and her supervisor Hermann Aberle, Professor in the Department of Functional Cell Morphology at Heinrich Heine University, Düsseldorf.
Hermann and Jaqueline
Hermann, can you give us your scientific biography and the questions your lab is trying to answer?
HA In retrospect, I find it quite interesting that my work was all the time somehow associated with cell adhesion and cellular junctions. I started off as a PhD student in Rolf Kemler‘s lab in Freiburg, working on vertebrate Cadherin-Catenin complexes, which connect neighbouring epithelial cells at adherens junctions. As a postdoctoral fellow, I switched topics and changed to Drosophila neuromuscular junctions (NMJs) in Corey Goodman‘s lab at UC Berkeley, where I was involved in a large-scale mutagenesis screen searching for genes that affected the morphology of these terminals. After cloning and functional characterization of the wishful thinking gene, I moved to Christiane Nüsslein-Volhard‘s lab at the MPI for Developmental Biology in Tübingen. There, we cloned several other genes that came out of the screen, including tolloid-related, mical, ankyrin-2 and neuroligin-1. During this time, and inspired by work of Darren Gilmour, we also developed tools and techniques to image migrating motor axons in living Drosophila embryos. Receiving a grant from the German Research Foundation and a generous invitation to join Christian Klämbt‘s department, we moved to the University of Münster, where we not only had fantastic imaging opportunities but also focussed more and more on the function of the axon guidance molecule sidestep (side), which also came out of the screen. In 2010, I received a call from the Heinrich Heine University in Düsseldorf, where we developed the idea to search for behavioural consequences of wiring defects.
Jaqueline, how did you come to join the lab, and what drives your research?
JK I studied biology at the University of Kassel and did my diploma thesis on the topic of spermatogenesis in Drosophila under the supervision of Mireille Schäfer. Afterwards, I wanted to continue my research with Drosophila, but I could imagine a change of topic and laboratory for my PhD very well. Hermann’s job advertisement and also his project description during my interview fascinated me very much. On the one hand, the correct wiring of the muscles is a multifaceted process in which many different components interlock and, on the other hand, the different methods and starting points spoke in favour of joining Hermann’s laboratory.
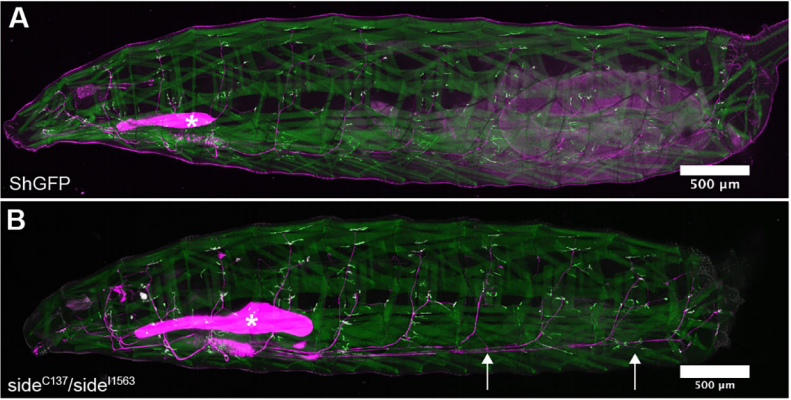
Control and side mutant larvae expressing the postsynaptic marker ShGFP and dsRed in motor acons and salivary glands. From Figure 1 in the paper.
When did your lab first get interested in the link between neuronal wiring and locomotion in flies?
HA Since the phenotype of side mutants is really strong, I was once asked after a talk if this has any consequences for viability or eclosion. This question stuck in my mind and I started to realize that most papers in the field dealt with the guidance process itself, in embryos, and hardly anybody was exploring postembryonic stages, when the neuromuscular system is actually functional and constantly contracting. Together with the Drosophila community moving more and more into circuit analysis and behavioural studies, I asked myself which behaviour could be affected by our mutations. It took a while until I realized that locomotion and movements are the underlying basis for most, if not all, behaviours. However, it took a while until we had all the necessary technical equipment. In fact, everything truly started as the high-speed video camera arrived.
Can you give us the key results of the paper in a paragraph?
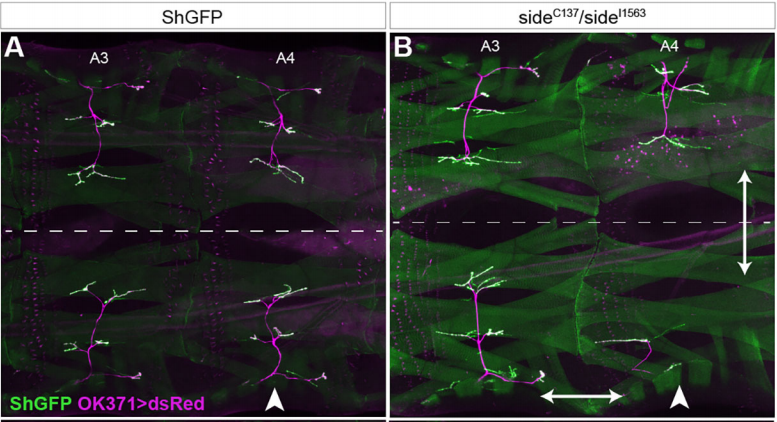
HA We examined the final innervation pattern on all muscles in side mutant third instar larvae. From this survey, we derived three major conclusions. First, innervation defects were permanent, i.e. even if potential rescue mechanisms exist, they failed to restore NMJs at non-innervated muscles. Second, loss of side affected all peripheral motor pathways and thus all body wall regions. Third, innervation errors were non-stereotypical and appeared in each hemisegment differentially. Since Side functions as a substrate-bound attractant, ventral bypass phenotypes or lack of NMJs at distal-most muscles, could be explained by insufficient attraction, which either inhibits defasciculation or slows axonal growth, respectively.
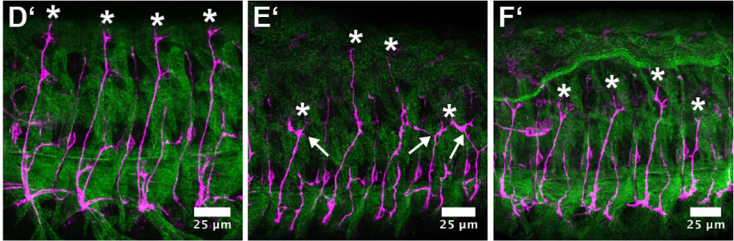
Overexpression of Side (E’) and Side-Cherry (F’) in muscle progenirots leads to premature stalling of motor axons, from Figure 3 in the paper
JK Also, overexpression of Side attracted motor axons at the wrong time to the wrong place, resulted in innervation defects. This was particularly evident when we expressed Side in muscle precursors. Attraction was that strong that individual axons travelling in the ISN were stretched into opposite directions leading to split pathways and complete lack of NMJs on dorsal-most muscles. This caused also amazing locomotion phenotypes. During crawling, larvae detached from the substrate and excessively lifted their head and tail segments into the air, despite lack of dorsal innervation. Nobody was expecting such a phenotype. Crawling seems to be much more complex than anticipated.
A dorsal view of third instar larvae showing some of the variability in the side phenotype, from Figure 2 in the paper.
side mutants show considerably variable phenotypes, even in adjacent hemisegments. What do you think this tells us about how axon guidance works in flies?
HA At the beginning of the screen, in Corey Goodman’s lab, we discussed a lot what kind of phenotypes we might possibly find. One such idea was that we might find genes that affect only a single NMJ on a specific muscle fibre, because we imagined a key-and-lock mechanism for wiring 30 muscle fibres, where unique ligands on motor axons connect to cognate receptors on muscles. Inactivating such a receptor by point mutations should in theory prevent the formation of the associated NMJ. Unfortunately, we did not find this phenotypic class, nor did anybody else. We therefore think that motor axon guidance in Drosophila is not hard-wired by high-affinity ligand-receptor complexes but rather functions via co-operative actions of several guidance molecules. Newer finding in Pablo Labrador‘s laboratory also push this older idea originally developed in the Keshishian and Goodman labs. Guidance decisions therefore seem to be made at several points and sum up along the entire path. If several decisions go wrong in a row, phenotypes become noticeable.
Do the locomotion defects you found in larvae have anything to tell us about movement disorders in mammals?
HA Oh yes, I think this is one of the interesting parts of our story. Similar to Drosophila, there are only a few mammalian studies that correlate axon guidance errors with muscle innervation patterns. But these studies made several interesting observations. First, in mice, if a motor nerve branch fails to develop or is severely stalled, entire muscle fields are not innervated after birth causing muscle atrophy. In limbs, this can lead to paralysis and aberrant locomotion patterns. Second, the phenotypes are not necessarily symmetrical and occasionally affect only one side of the body, i.e. very similar to the unilateral defects observed in side mutants. Third, feet and ankles of affected limbs are occasionally twisted inward in newborn mice, a deformity highly reminiscent to congenital clubfoot in humans. The aetiology of clubfoot is still not fully understood, but since it frequently develops unilaterally, one speculative possibility is that it could develop due to innervation defects.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
JK There was not one key moment that has stuck with me, but several small highlights, especially at the laser-scanning microscope. I am over and over again enthusiastic about how “beautiful” the axonal pattern in the embryo or larva is. Further, I had such special moments when larvae showed particularly strong or unusual phenotypes during crawling, which looks sometimes quite funny.
Drosophila larva overexpressing Side in muscles and showing an extreme crawling phenotype.
And what about the flipside: any moments of frustration or despair?
JK Yes, there were some of these frustrating moments, especially handling living larvae for videography. During my research, I have realized that Drosophila larvae have sometimes their own mind, as they decided not to crawl at all or not to crawl in a straight line for the high-speed videography, but rather to crawl away from the nicely prepared agar block.
What next for you after this paper?
JK Since this paper is only a partial project of my PhD thesis, I plan for the next months to press ahead with another project for publication – hopefully we can submit the manuscript successfully at the end of this year.
And where will this work take the Aberle lab?
HA One direction we are heading is how locomotion in animals with hydrostatic skeletons works after all. Crawling behaviour is much more sophisticated and fine-tuned than it appears at first glance. We would like to understand how specific muscle groups contribute to particular movements and how this is antagonized by the liquid-filled body cavity. Which muscle groups, for example, induce rolling escape behaviours during attacks of predatory wasps or coordinate jumping in some dipteran larvae. Ultimately we would like to find mutations or conditions which activate or inactivate particular movements, in order to better understand the underlying circuitry and machinery. Another line of research will certainly be the functional analysis of the entire Sidestep family. There are 7 Side paralogs in the Drosophila genome and virtually nothing is currently known about their functions.
The Düsseldorf skyline. Image credit: Ingo Valentin, from Wikipedia.
Finally, let’s move outside the lab – what do you like to do in your spare time in Düsseldorf?
JK As I am a very water-loving person, I like to spend my spare time along the Rhine river – whether for a walk or to make myself comfortable on a blanket on its shores and let my soul dangle watching the ships passing by. I also like to take advantage of the city’s concert events, as two of my favourite bands, “Die Toten Hosen” and “Broilers”, come from Düsseldorf and regularly give concerts in the city. Furthermore, the whole laboratory and I like to celebrate the Düsseldorf Carnival, which takes place every spring.
HA Düsseldorf has a quite lively art scene, and since I enjoyed landscape photography very much during my undergraduate years, I am particularly drawn to the “Düsseldorf School of Photography”, founded by Bernd and Hilla Becher. Whenever there are exhibitions by Andreas Gursky, Axel Hütte, Thomas Ruff, to name a few, I try to not miss the vernissage. I also love to shop for coffee-table books in local stores.
Welcome to our monthly trawl for developmental biology (and other related/just plain cool) preprints.
This month we found a tonne of papers dealing with various aspects of inheritance in worms, a flush of fly mechanics, and more single cell sequencing than you could shake a stick at. And as summer draws to a close, it’s raining cats and dogs (and wolves) in our ‘Why not…’ section.
The preprints were hosted on bioRxiv, PeerJ, andarXiv. Let us know if we missed anything, and use these links to get to the section you want:
Conserved cell types with divergent features between human and mouse cortex
Rebecca D Hodge, Trygve E Bakken, Jeremy A Miller, Kimberly A Smith, Eliza R Barkan, Lucas T Graybuck, Jennie L Close, Brian Long, Osnat Penn, Zizhen Yao, Jeroen Eggermont, Thomas Hollt, Boaz P Levi, Soraya I Shehata, Brian Aevermann, Allison Beller, Darren Bertagnolli, Krissy Brouner, Tamara Casper, Charles Cobbs, Rachel Dalley, Nick Dee, Song-Lin Ding, Richard G Ellenbogen, Olivia Fong, Emma Garren, Jeff Goldy, Ryder P Gwinn, Daniel Hirschstein, C Dirk Keene, Mohamed Keshk, Andrew L Ko, Kanan Lathia, Ahmed Mahfouz, Zoe Maltzer, Medea McGraw, Thuc Nghi Nguyen, Julie Nyhus, Jeffrey G Ojemann, Aaron Oldre, Sheana Parry, Shannon Reynolds, Christine Rimorin, Nadiya V Shapovalova, Saroja Somasundaram, Aaron Szafer, Elliot R Thomsen, Michael Tieu, Richard H Scheuermann, Rafael Yuste, Susan M Sunkin, Boudewijn Lelieveldt, David Feng, Lydia Ng, Amy Bernard, Michael Hawrylycz, John Phillips, Bosiljka Tasic, Hongkui Zeng, Allan R Jones, Christof Koch, Ed S Lein
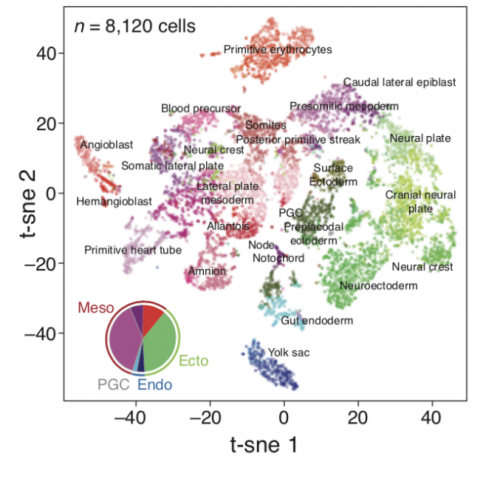
t-sne plot of an E8.5 mouse embryo, from Chan, et al.’s preprint
Molecular recording of mammalian embryogenesis
Michelle Chan, Zachary D Smith, Stefanie Grosswendt, Helene Kretzmer, Thomas Norman, Britt Adamson, Marco Jost, Jeffrey J Quinn, Dian Yang, Alexander Meissner, Jonathan S Weissman
Planar cell polarity pathway and development of the human visual cortex
Jean Shin, Shaojie Ma, Edith Hofer, Yash Patel, Gennady Roshchupkin, Andre M Sousa, Xueqiu Jian, Rebecca Gottesmann, Thomas H Mosley, Myriam Fornage, Yasaman Saba, Lukas Pirpamer, Reinhold Schmidt, Helena Schmidt, Bernard Mazoyer, Amaia Carrion-Castillo, Joshua Bis, Shuo Li, Qiong Yang, Michelle Luciano, Sherif Karama, Lindsay Lewis, Mark Bastin, Matthew A Harris, Ian Deary, Joanna M Wardlaw, Markus Scholz, Markus Loeffler, Veronica Witte, Frauke Beyer, Arno Villringer, Hieab HHH Adams, M Arfan Ikrum, William S Kremen, Nathan A Gillespie, Nenad Sestan, Zdenka Pausova, Sudha Seshadri, Tomas Paus
Bovine ovary sections in Plewes, et al.’s preprint
Mapping cell types in the foetal human neocortex, from Polioudakis, et al.’s preprint
A single cell transcriptomic analysis of human neocortical development
Damon Polioudakis, Luis de la Torre-Ubieta, Justin Langerman, Andrew G Elkins, Jason L Stein, Celine K Vuong, Carli K Opland, Daning Lu, William Connell, Elizabeth K Ruzzo, Jennifer K Lowe, Tarik Hadzic, Flora I Hinz, Shan Sabri, William E Lowry, Kathrin Plath, Daniel H Geschwind
Epigenetic factors coordinate intestinal development
Julia Ganz, Ellie Melancon, Catherine Wilson, Angel Amores, Peter Batzel, Marie Strader, Ingo Braasch, Parham Diba, Julie A Kuhlman, John H Postlethwait, Judith S Eisen
Analysis of novel domain-specific mutations in the zebrafish ndr2/cyclops gene generated using CRISPR-Cas9 RNPs
Ashley N Turner, Reagan S Andersen, Ivy E Bookout, Lauren N Brashear, James C Davis, David M Gahan, John P Gotham, Baraa A Hijaz, Ashish S Kaushik, Jordan B McGill, Victoria L Miller, Zachariah P Moseley, Cerissa L Nowell, Riddhi K Patel, Mia C Rodgers, Yazen A Shihab, Austin P Walker, Sarah R Glover, Samantha D Foster, Anil Kumar Challa
Functional dissection of the ARGONAUTE7 promoter
J Steen Hoyer, Jose L Pruneda-Paz, Ghislain Breton, Mariah A Hassert, Emily E Holcomb, Halley Fowler, Kaylyn M Bauer, Jacob Mreen, Steve A Kay, James C Carrington
Gene Correction for SCID-X1 in Long-Term Hematopoietic Stem Cells
Mara Pavel-Dinu, Volker Wiebking, Beruh T Dejene, Waracharee Srifa, Sruthi Mantri, Carmencita Nicolas, Ciaran Lee, Gang Bao, Eric J Kildebeck, Niraj Punjya, Camille Sindhu, Matthew A Inlay, Nivi Saxena, Suk See DeRavin, Harry Malech, Maria Grazia Roncarolo, Kenneth I Weinberg, Matthew Porteus
Neural crest stem cells from Stebbins, et al.’s preprint
Signalling pathways drive heterogeneity of ground state pluripotency
Kirsten R McEwen, Sarah Linnett, Harry G Leitch, Prashant Srivastava, Lara Al-Zouabi, Tien-Chi Huang, Maxime Rotival, Alex Sardini, Thalia E Chan, Sarah Filippi, Michael Stumpf, Enrico Petretto, Petra Hajkova
Need for high-resolution Genetic Analysis in iPSC: Results and Lessons from the ForIPS Consortium
Bernt Popp, Mandy Krumbiegel, Janina Grosch, Annika Sommer, Steffen Uebe, Zacharias Kohl, Sonja Ploetz, Michaela Farrell, Udo Trautmann, Cornelia Kraus, Arif B Ekici, Reza Asadollahi, Martin Regensburger, Katharina Guenther, Anita Rauch, Frank Edenhofer, Juergen Winkler, Beate Winner, Andre Reis
A human cell model of cardiac pathophysiological valvulogenesis
Tui Neri, Emylie Hiriart, Piet Van Vliet, Emilie Faure, Russel Norris, Batoul Farhat, Julie Lefrancois, Thomas Moore-Morris, Stephane Zaffran, Randolph Faustino, Alexander Zambon, Jean-Pierre Devisgnes, David Salgado, Yukiko Sugi, Robert Levine, Jose Luis de la Pompa, Andre Terzic, Sylvia Evans, Roger Markwald, michel Puceat
Caenorhabditis uteleia under SEM, from Stevens, et al.’s preprint
Comparative genomics of ten new Caenorhabditis species
Lewis Stevens, Marie-Anne Félix, Toni Beltran, Christian Braendle, Carlos Caurcel, Sarah Fausett, David HA Fitch, Lise Frézal, Taniya Kaur, Karin C Kiontke, Matt D Newton, Luke M Noble, Aurélien Richaud, Matthew V Rockman, Walter Sudhaus, Mark Blaxter
The Genomic Basis of Arthropod Diversity
Gregg W. C. Thomas, Elias Dohmen, Daniel S. T. Hughes, Shwetha C. Murali, Monica Poelchau, Karl Glastad, Clare A. Anstead, Nadia A. Ayoub, Phillip Batterham, Michelle Bellair, Gretta J Binford, Hsu Chao, Yolanda H Chen, Christopher Childers, Huyen Dinh, HarshaVardhan Doddapaneni, Jian J Duan, Shannon Dugan, Lauren A Esposito, Markus Friedrich, Jessica Garb, Robin B. B Gasser, Michael A. D. Goodisman, Dawn E Gundersen-Rindal, Yi Han, Alfred M Handler, Masatsugu Hatakeyama, Lars Hering, Wayne B Hunter, Panagiotis Ioannidis, Joy C Jayaseelan, Divya Kalra, Abderrahman Khila, Pasi K Korhonen, Carol Eunmi Lee, Sandra L Lee, Yiyuan Li, Amelia R.I. Lindsey, Georg Mayer, Alistair P McGregor, Duane D. McKenna, Bernhard Misof, Mala Munidasa, Monica Munoz-Torres, Donna M Muzny, Oliver Niehuis, Nkechinyere Osuji-Lacy, Subba R. Palli, Kristen A. Panfilio, Matthias Pechmann, Trent Perry, Ralph S. Peters, Helen C Poynton, Nikola-Michael Prpic, Jiaxin Qu, Dorith Rotenberg, Coby Schal, Sean D Schoville, Erin D Scully, Evette Skinner, Daniel B Sloan, Richard Stouthamer, Michael R Strand, Nikolaus U Szucsich, Asela Wijeratne, Neil D Young, Eduardo E Zattara, Joshua B Benoit, Evgeny M Zdobnov, Michael E Pfrender, Kevin J Hackett, John H Werren, Kim C Worley, Richard A Gibbs, Ariel D Chipman, Robert M Waterhouse, Erich Bornberg-Bauer, Matthew W Hahn, Stephen Richards
Spatial Organization of Rho GTPase signaling by RhoGEF/RhoGAP proteins
Paul Markus Mueller, Juliane Rademacher, Richard D Bagshaw, Keziban Merve Alp, Girolamo Giudice, Louise E Heinrich, Carolin Barth, Rebecca L Eccles, Marta Sanchez-Castro, Lennart Brandenburg, Geraldine Mbamalu, Monika Tucholska, Lisa Spatt, Celina Wortmann, Maciej T Czajkowski, Robert William Welke, Sunqu Zhang, Vivian Nguyen, Trendelina Rrustemi, Philipp Trnka, Kiara Freitag, Brett Larsen, Oliver Popp, Philipp Mertins, Chris Bakal, Anne-Claude Gingras, Olivier Pertz, Frederick P Roth, Karen Colwill, Tony Pawson, Evangelia Petsalaki, Oliver Rocks
The Signaling Pathways Project: an integrated ‘omics knowledgebase for mammalian cellular signaling pathways
Scott Ochsner, David Abraham, Kirt Martin, Wei Ding, Apollo McOwiti, Zichen Wang, Kaitlyn Andreano, Ross Hamilton, Yue Chen, Angelica Hamilton, Marin Gantner, Michael Dehart, Shijing Qu, Susan Hilsenbeck, Lauren Becnel, Dave Bridges, Avi Maayan, Janice Huss, Fabio Stossi, Charles Foulds, Anastasia Kralli, Donald McDonnell, Neil McKenna
The intent of this book is to demonstrate the essential role of cellular mechanisms in transforming the genotype into the phenotype by transforming gene activity into evolutionary change in morphology. This book evaluates the evolution of cells themselves and the role cells play as agents of change at other levels of biological organization. Chapters explore Darwin’s use of cells in his theory of evolution as well as Weismann’s theory of the separation of germ plasm from body cells that influenced our understanding that acquired changes are not passed on to future generations. The study of evolution through the analysis of cell lineages during embryonic development is discussed. Discovery that cells exchange organelles via symbiosis led to a fundamental reevaluation of prokaryotic and eukaryotic cells and to a reorganization of the Tree of Life. Identification of cellular signaling centers and mechanisms responsible for cellular patterning as mediators of phenotypic change during evolution is presented. Chapters present evidence for the powerful new synergies between cell biology and evolutionary theory.
The Company of Biologists’ journals – Development, Journal of Cell Science, Journal of Experimental Biology andDisease Models & Mechanisms – offer Travelling Fellowships of up to £2,500 or currency equivalent to graduate students and post-doctoral researchers wishing to make collaborative visits to other laboratories. These are designed to offset the cost of travel and other expenses. There is no restriction on nationality.
The deadline for the current round of applications is tomorrow – 31 August!
Have you heard of an animal that can lose most of its body tissues and the remnant tissues aggregate to regenerate the lost parts and recovery its original form?
Do you know an animal that can quickly colonize marine surfaces by asexual reproduction, just like weed would in terrestrial environments ?
Do you know an animal that can disperse to new locations when small portions of its body separate and move away from the rest?
These descriptions may remind you of characters from science fiction stories, but these are real characteristics of colonial animals. Animals colonies are composed of discrete multicellular units (e.g zooids, polyps), that are physiologically interconnected and undergo clonal replication maintaining an identical genotype throughout all of their components (Hughes, 1989; Jackson & Coates, 1986) . This includes a number of marine and freshwater animals, such as many corals and hydroids, bryozoans, some hemichordates and some ascidians (a group within the subphylum Tunicata). One of the main projects in the laboratory is to study the evolution of colonial life strategyin ascidians, in which coloniality evolve several times.
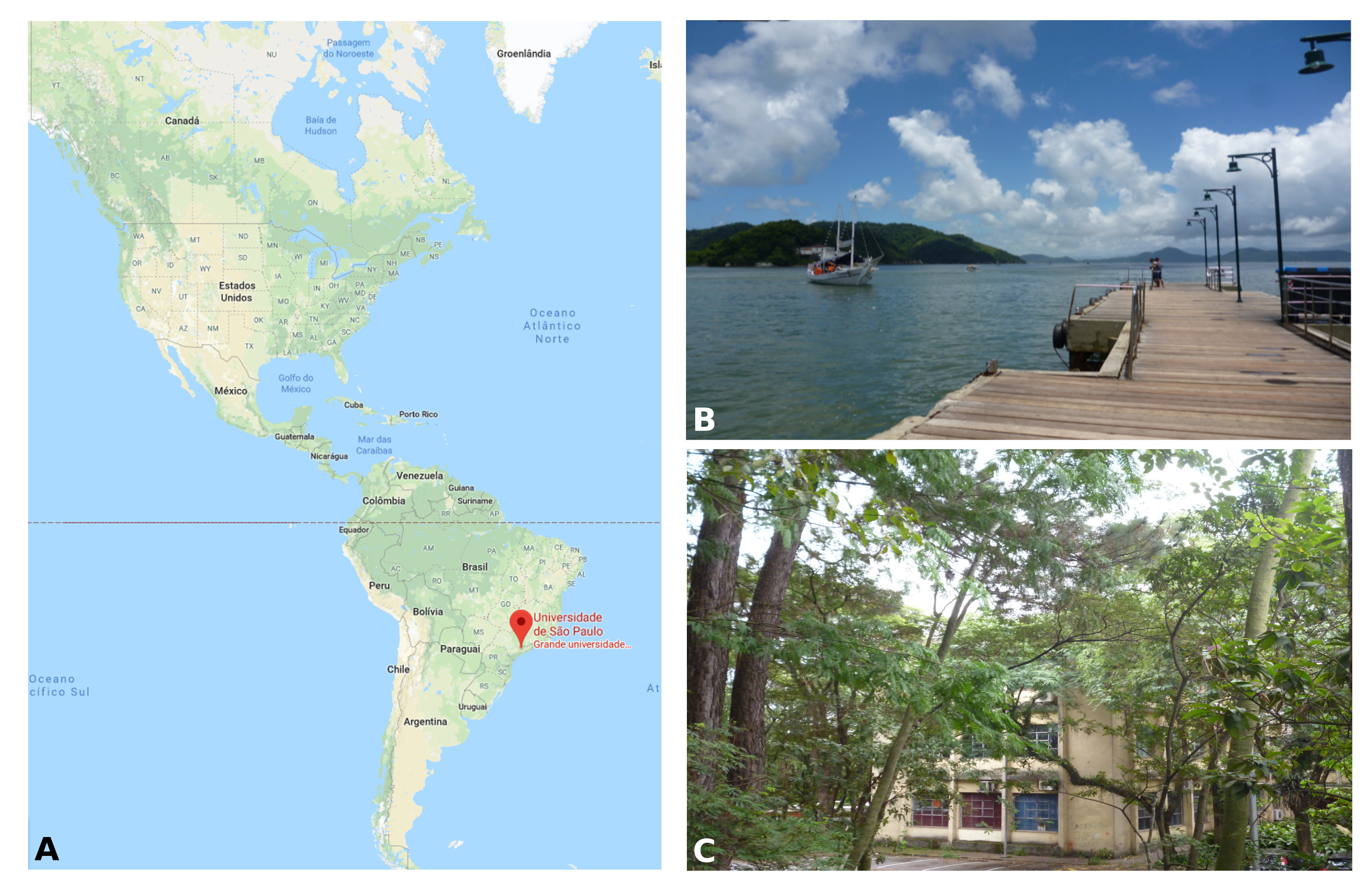
I am a PhD student in Prof. Federico Brown’s laboratory (http://zoologia.ib.usp.br/evodevo2/) in the Department of Zoology at the University of São Paulo. We are located in a subtropical area in the south of Brazil, in a huge metropolis 100 km away from the ocean. In fact, our lab is located next to a small forest reserve on campus that is reminiscent of the Mata Atlantica Forest (a hotspot of biodiversity), from the laboratory’s windows, we can see a great diversity of birds, insects, spiders and sometimes even monkeys (Fig.1).
Fig. 1 Location of the Federico Brown’s laboratory and ascidian collection sites. (A) Location of the University of São Paulo. (B) A port in Santos city where we collect and grow the colonial tunicates. (C) View of the laboratory at the University of São Paulo.
Our work in the laboratory begins with the collection of tunicates from the ocean. We travel by car to close coastal cities, including São Sebastião (4h away) and Santos (1h away) (Fig. 1B). We collect the tunicates from pilings and floats in the port marinas. The colonial species are carefully collected and attached to glass slides using thread (Fig. 2A). The slides are placed in slide boxes, with open windows on both sides to allow water circulation, and the boxes are hung to the dock with ropes. After two weeks, the colonies grow to cover the slides. Therefore attached tunicates are cleaned, transported to the aquarium system, and are ready for use in experiments (Fig. 2).
Fig. 2 Procedures to attach and cultivate ascidians. (A) Colonial ascidians are attached with a thread to a glass slide. (B) Colonial ascidians on the glass slides are placed in racks for culturing. (C) At our field sites, glass slides with ascidians are placed in slide boxes with windows and hung from the dock to allow them to grow in the ocean
For my PhD project, I am working with the genus Symplegma, a member of the family Styelidae, in which coloniality arose at least twice from solitary ancestors. Symplegma is the sister genus to the botryllids (Botryllus and Botrylloides), a group of highly integrated colonial species that undergo weekly cycles of asexual development (Brown et al., 2009). Symplegma has less integrated colonies and characteristics more similar to the solitary species. Thus it is an interesting genus to study the evolution of coloniality.
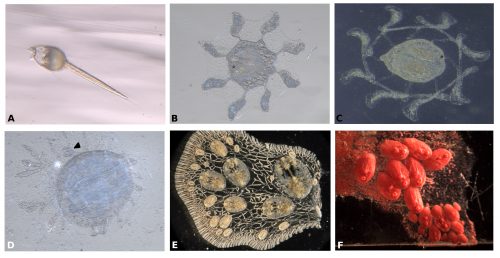
Symplegma, as other colonial ascidians, has internal fertilization and brooding, with embryos that are incubated for several days to weeks before the tadpole larvae are released. The tadpole larva have a brief period of swimming and settle rapidly (Fig. 3A). During settlement, the larvae undergo metamorphosis, in which most of the tail structures are resorbed and the mouth rotates close to 90º to the dorsal side, changing from a tailed swimming larva into a sessile filter feeder (Fig. 3).
Fig. 3 Stages of Symplegma development. (A) Newly released larva of the species Symplegma rubra. (B) Settled larva in the process of metamorphosis, with extended ampullae. (C) Later stage of metamorphosis, where the primary ampullae have fused. (D) First zooid and bud (arrowhead) formed after metamorphosis. (E) Symplegma brakenhielmi colony. (F) Symplegma rubra colony.
During metamorphosis, the ampullae extending a symmetrical pattern around the first zooid (Fig. 3B). The ampullae are peripheral pouch-like structures of the blood vessel system, essential for communication of the colony with its environment. Then ampullae continue to extend and, interconnect to form the primary net of blood vessels, in which the blood circulate constantly (Fig. 3C). Then this system of vessels grows forming more vessels that connect the zooid and the first bud (Fig. 3D).
Inside the vessels, specialized blood cells circulate, constantly coordinating biological processes between the zooids of the colony (Video 1).
For example, the circulatory cells are key to the clonal formation of new buds and the coordination of death of old zooids. New buds can be formed in two locations, either at the lateral epithelium of the adult zooid or along blood vessels that are far from adult zooids (Fig. 3E-F).
While some colonial styelids have highly integrated colonies with coordinated cycles of degeneration and regeneration of zooids, Symplegma is much less integrated and coordinated. We are interested in how developmental and regenerative processes differ between species with different degrees of coordination and integration. For example, we conduct experiments to observe whole body regeneration by removing all the zooids and buds retaining only the blood vessels of the original colony. Immediately after surgery, the blood coagulates and circulation in the remnant vessels stops. Twelve hours after the surgery, blood circulation is restored without any zooids or heart to pump the blood. Presumably, this circulation is caused by vessel contractions (Video 2).
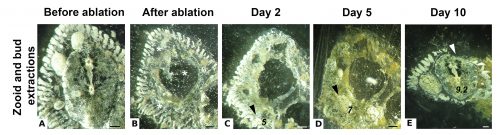
Next, the remaining vascular tissue aggregates and form a mass, in which new zooids arise. Ten days after surgery a complete functional colony has regenerated (Fig.4).
Fig. 3 Regeneration in a Symplegma brakenhielmi colony. (A) Colony before ablation, in which a fully differentiated zooid is shown. (B) Colony immediately after ablation, zooid and buds were removed (asterisks) and only the vascular tissues remain. (C) Day 2 after ablation, vascular tissues have aggregated and new buds are observed at stage 5 (arrowhead); asterisk shows the scar of the surgery. (D) Day 5 after ablation, the vascular tissues form a mass and a bud (arrowhead) continues to develop (stage 7). (E) Day 10 after ablation, the new zooid is fully differentiated (stage 9.2). Scale bar is 500 µm. (Edited from Gutierrez & Brown, 2017).
These results suggest that the vascular tissue has the capacity to rearrange itself and regenerate new zooids. Our results show that Symplegma colonies act like self-regulating systems that have the ability to rearrange its components (blood vessels and blood cells) after perturbations to regenerate damaged or lost parts. Due to the presence of replaceable zooids, the colonial life history allows for the recovery of lost parts by regeneration, fast colonization of marine substrates, and high survival rates after predation or weather-related disturbances. These colonial animals are like superorganisms!! They show amazing developmental mechanisms linked to coloniality.
If you would like any more information about their life history and how to work on them, just ask us in the comments or via email [as.gutierrez57@ib.usp.br].
References
-Brown, F. D., Tiozzo, S., Roux, M. M., Ishizuka, K., Swalla, B. J., & De Tomaso, A. W. (2009). Early lineage specification of long-lived germline precursors in the colonial ascidian Botryllus schlosseri. Development (Cambridge, England), 136(20), 3485–3494.https://doi.org/10.1242/dev.037754
-Gutierrez, S., & Brown, F. D. (2017). Vascular budding in Symplegma brakenhielmi and the evolution of coloniality in styelid ascidians. Developmental Biology, 423(2). https://doi.org/10.1016/j.ydbio.2017.01.012
-Hughes, R. (1989). A Functional Biology of Clonal Animals. New York: Chapman and Hall.
-Jackson, J. B. C., & Coates, a. G. (1986). Life Cycles and Evolution of Clonal (Modular) Animals. Philosophical Transactions of the Royal Society B: Biological Sciences, 313(1159), 7–22. https://doi.org/10.1098/rstb.1986.0022
Very excited to invite applications from post-doctoral researchers to join my lab to work on a Leverhulme Trust- funded project to look at the mechanisms regulating branching in Selaginella kraussiana. I would like to use a candidate gene approach, looking at Selaginella PIN and TCP function. I have written a bit about the project here, and you can apply here.
When people digest food, the flow of fluids and digestive enzymes to the gut is critical. Until recently, researchers had marvelled at the incredibly complex system of ducts that transports a stream of enzymes and mucus from the pancreas to the gut. Astonishingly, a research project led by Professor Grapin-Botton has revealed that the ducts are created similar to river beds. This new knowledge may lead to improving treatment for people with cystic fibrosis and diabetes.
Most people associate transport with roads, rivers or rails. In nature, transport is equally vital. Trees have a transport system in their roots, branches and leaves, and humans have many transport channels such as nerve fibres and lung bronchioles. Many of these structures are nearly identical from person to person. However, this does not apply to the transport ducts between the pancreas and the gut, which are essential for efficient digestion. A Danish research project has now solved the enigma of how the ducts are created to transport mucus, enzymes and chemical substances.
“The transport channels from the pancreas to the gut are critical for digesting food and neutralizing acidic gastric juices. Because efficient transport is essential, we wondered why the ducts vary from person to person. Our new results show that the channels can change similarly to a river bed during fetal life. The channels with the greatest flow appear to widen, whereas others run dry and disappear. This knowledge may help us to treat people with cystic fibrosis and some forms of monogenic diabetes associated with cystic ducts, who have such transport problems,” explains Anne Grapin-Botton,
A surprising result
To map how the ducts inside the pancreas are formed, the researchers marked the ducts with fluorescent antibodies, which enabled them to see when a duct was formed and its connection to others. The researchers monitored the development of the pancreas in mice.
“During organ development in fetuses, the ducts are initiated from small holes. These small holes connect and fuse together, thereby creating many ducts that develop into a complex network. It resembles a town with a labyrinth of streets. What fascinated us was how, from this labyrinth, a simpler treelike structure emerged at birth.”
To keep track of the high volume of data and to identify how the network of ducts changed, the researchers teamed-up with Kim Sneppen, a professor and physicist from the Niels Bohr Institute, University of Copenhagen. Svend Bertel Dahl-Jensen, their PhD student, pulled together the threads and uncovered the secrets of the labyrinth.
“We decided to adapt the programs used to model road, rail or internet networks, and the picture that emerged once we inputted the data into the computer reminded us of something familiar: a river system. And indeed, going back to the laboratory, we found that as soon as the ducts were formed they secreted juice. We think that this creates a flow of fluid towards the intestine already in fetuses. Some ducts widen, while others run dry and disappear.”
Pancreatic ducts (as well as the surrounding duodenum) are shown in green (stained for β-catenin) and the islets of Langerhans appear in red (stained for insulin). Image Credit: Dror Sever
Ducts collapse
The study showed that the cells from the collapsed ducts did not simply disappear. They are likely recycled to widen existing ducts. This may turn out to be important for medicine.
“People with cystic fibrosis or with certain forms of monogenic diabetes have problems in the pancreatic ducts. For diabetes, we do not know why diabetes and enlarged ducts are associated when certain genes are mutated.”
People with cystic fibrosis have defective pancreatic ducts. This results from mutations in a gene that codes for a channel enabling fluid secretion inside the ducts. The researchers demonstrated that the secretion defects may start very early in fetuses and their work may lead them to consider earlier treatment.
“We would now really like to understand why people whose pancreatic ducts have collapsed are more likely to develop diabetes. One hypothesis we are pursuing is that they make less beta cells secreting insulin. These cells are initially formed in the ducts. We are now studying this in mouse models and in miniature human pancreas (organoids) made from stem cells in 3D culture.”
In our recently published paper https://elifesciences.org/articles/34880, we report that the transcription factor Pitx2c has an unexpected role during gastrulation, where it acts cell non-autonomously to promote mesendodermal cell migration required for axis extension in zebrafish.
“It is not birth, marriage or death which is the most important time in your life, but gastrulation.
– Lewis Wolpert, 1989
Undoubtedly, gastrulation is a critical time during development, as the entire body plan is defined by what happens at this stage. Cells must undergo intricate and complex movements to generate the three germ layers and end up in the correct location to give rise to all of the parts of the body. Not only is it an absolutely beautiful event to observe, it is also an essential developmental process that is well-suited for probing important questions surrounding cell behaviour during migration and embryonic patterning. This process is especially clear in the zebrafish embryo – its optical transparency and amenability to live imaging allow one to attain single-cell resolution to examine cellular behaviours in real-time.
How it all started…
When I joined Didier Stainier’s lab as a postdoc, I wanted to focus on questions surrounding cardiac laterality in zebrafish. While it has been known for over 20 years that the transcription factor Pitx2c is a critical player in defining the left side of the body, many questions surrounding how Pitx2c directs this process remain to be explored. Around this time, Didier was in contact with Patrick Blader from the Centre de Biologie du Développement, Université Paul Sabatier in Toulouse. Patrick’s lab had recently generatedzebrafish mutant alleles in pitx2, and he generously shared his line with us.
We started analysing these mutants, and at first, I was rather disappointed that we did not see any defects in cardiac laterality! However, when we looked at the phenotypes in embryos lacking both maternal and zygotic (MZ) Pitx2 function, we observed many embryos at 24 hours post-fertilization with shorter bodies and somite defects. These phenotypes were reminiscent of many of the gastrulation mutants recovered in ENU screens, and so we began exploring the role for maternal and zygotic Pitx2c function in more detail.
What we learned…
We first questioned whether any patterning defects were present in MZpitx2c mutants during gastrulation and early somitogenesis and looked at the expression of many genes by in situ hybridization. While the different germ layers were present, it was obvious that the cells were not in their correct location. For example, we observed that the cells giving rise to the notochord were located in a wider domain than in wild-type embryos. We also found that at late gastrulation, mesendodermal cells looked rather disorganized, arguing that cell migration was likely affected in these mutants.
Gastrulation movements in zebrafish have been very well described. While epiboly spreads the cells of the epiblast down around the yolk, mesendodermal precursors separate from the epiblast and migrate away from the margin via internalization movements. Internalization is followed by convergence and extension movements that mediolaterally narrow the embryo, while elongating them along the anterior-posterior axis. We first focused on assessing convergence and extension movements in the mutants since they presented phenotypes suggestive of defects in these cell behaviours.
To test whether convergence and extension movements were affected in the MZpitx2c mutants, we photoconverted lateral mesendodermal cells and tracked their migration over time. These experiments suggested that both dorsal convergence and anterior extension were reduced. Similarly, we observed that the distance between the prechordal plate and the notochord at the 1 somite stage was reduced in MZpitx2c mutants compared to wild types, and when we looked at cell morphology within the notochord of mutants, we found that the cell shapes were rounder and failed to elongate mediolaterally as in wild-type embryos. These data suggested to us that Pitx2c functions to promote convergence and extension movements.
Previous work from our group and others has described the movements of endodermal cells. In this population of cells, early gastrulation movements undergo a ‘random walk’ motion to spread endodermal cells across the embryo; at late gastrulation, endodermal cells rapidly increase their speed and migration trajectories, becoming straighter and oriented towards the midline to form the endodermal sheet. When we examined these behaviours in MZpitx2c mutants, we observed that the transition between random walk motion to oriented, persistent migration was blunted. Taken together, our data indicate that cell migration behaviours during late gastrulation are strongly disrupted in the absence of maternal and zygotic Pitx2c function.
We then performed transplantation assays, which really gave us critical insight into the role of Pitx2c. In these experiments, we transplanted mesendodermal cells that expressed LIFEACT-GFP into unlabelled host embryos, allowing us to analyse cell migration behaviours at single-cell resolution. What was especially striking were the observations that wild-type cells that were transplanted into MZpitx2c hosts sent out fewer projections and their morphology was flattened. These key observations indicated that Pitx2 is required cell non-autonomously to promote migration behaviours of mesendodermal cells.
Movie: WT cells expressing LIFEACT-GFP were transplanted into WT (left panel) or MZpitx2c mutant (right panel) hosts. Transplanted WT cells exhibit increased surface area and extend fewer protrusions that appear more actin-dense in mutant hosts compared to WT hosts.
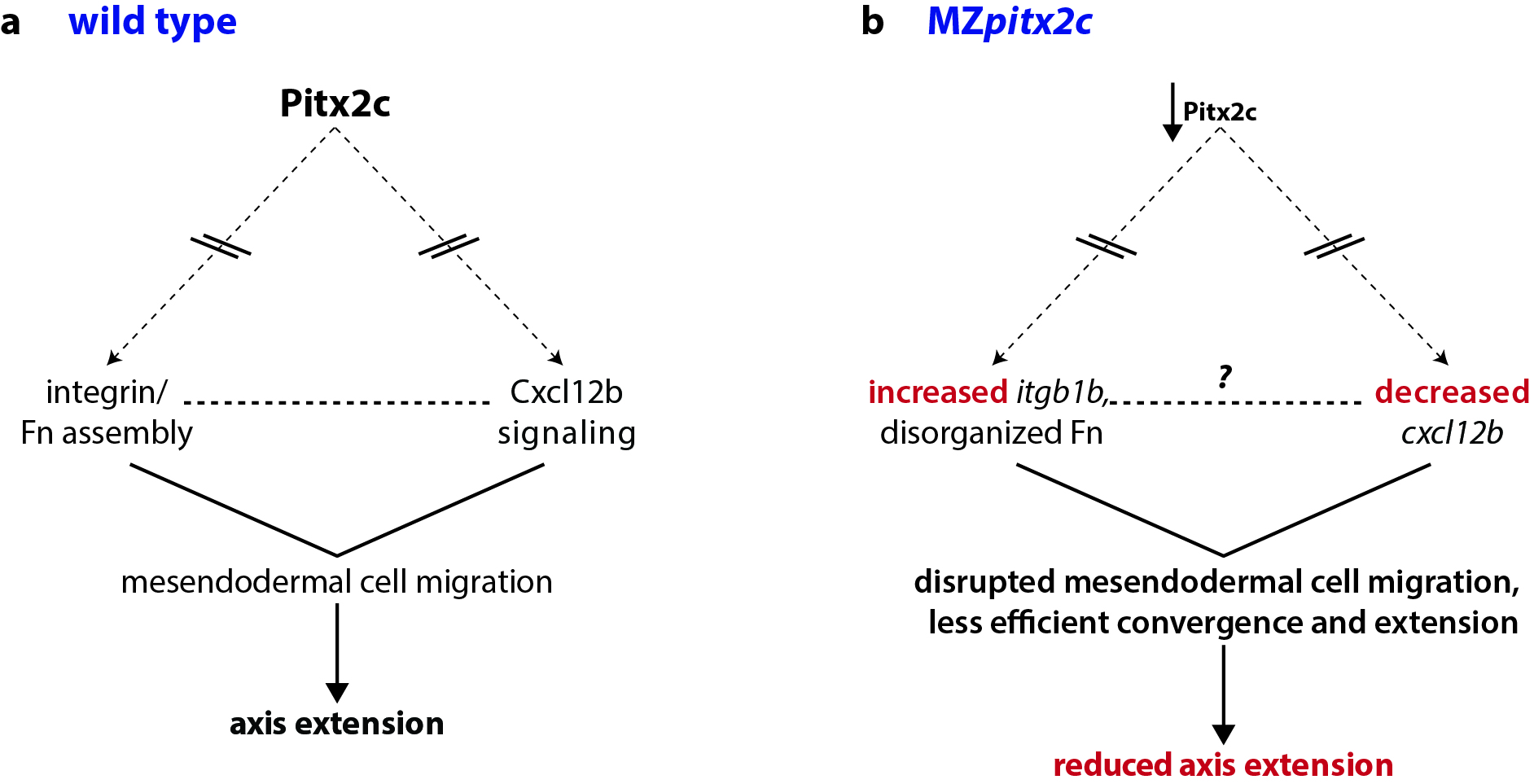
At the same time, we were also analyzing microarray data from MZpitx2c mutants and in embryos where we had injected pitx2c mRNA at the 1-cell stage. One of the top hits in these analyses was a gene encoding the chemokine ligand Cxcl12b. A few years ago, the Schilling and Kikuchi labs showed that mesodermal expression of cxcl12b (or sdf1b) was important to coordinate and guide the movements of the endoderm, which express the receptor gene cxcr4a. They also proposed that Cxcl12b/Cxcr4a signaling promotes integrin-mediated adhesion for coordinated mesendodermal cell migration. Furthermore, Cxcl12b/Cxcr4a signaling regulates the formation of filopodial processes in gastrulating mesendodermal cells.
Together, these data fit very nicely with the phenotypes we had observed in MZpitx2c mutants, and so we focused our analyses of these pathways. Indeed, we found disrupted expression of cxcl12b and the integrin subunit gene itgb1b, as well as altered deposition of Fibronectin between mesendodermal cells. Therefore, these data suggested that Pitx2 functions upstream of chemokine-dependent changes in adhesion to the ECM to influence cell migration.
What does it mean?
Pitx2 is a transcription factor that plays diverse roles in many tissues and developmental contexts. In humans, mutations in PITX2 coding regions lead to Axenfeld-Rieger syndrome, while promoter/enhancer mutations are associated with atrial fibrillation. Despite these clinically relevant syndromes, few downstream pathways have been identified. In our paper, we describe a role for Pitx2c where it promotes convergence and extension movements during gastrulation. Using transcriptomic analyses of pitx2c gain- and loss-of-function embryos, we identify transcriptional changes in genes involved in chemokine-ECM interaction and signaling.
We hypothesize that the early function of Pitx2 during gastrulation is evolutionarily conserved. Recent work from the Stern and Viebahn groups has shown that Pitx2 expression precedes the formation of the primitive streak in chick and rabbit embryos. We were also very excited to find expression of pitx2 in single-cell RNAseq datasets of early gastrula stage mice and frogs, suggesting that Pitx2 is also playing an early role in these species.
These studies raise the question as to whether similar mechanisms downstream of Pitx2c are at play in other developmental contexts. As the Nodal-Lefty-Pitx2 cascade appears to be conserved at both the onset of gastrulation and during the establishment of left-right identity, it will be exciting to explore whether the downstream pathways we identify here are conserved in other contexts.
It’s an age-old mystery of the heart: do opposites attract, or will like do better with like? We can now answer this pressing question, at least for Drosophila cardioblasts: cells prefer to ‘swipe right’ on a shared transcriptional profile, but the resulting relationships are stronger if there are some unattractive alternatives around to remind them to love the one they’re with.
To put that in more scientific terms, in order to build complex structures that perform versatile functions, biological systems need to be able to specific and precise cell-cell connections. Yet the question of how cells find the right partners as organs form generally remains poorly understood. In our recently published work (Zhang et al., 2018), our team has uncovered some of the secrets of how cells make the perfect match.
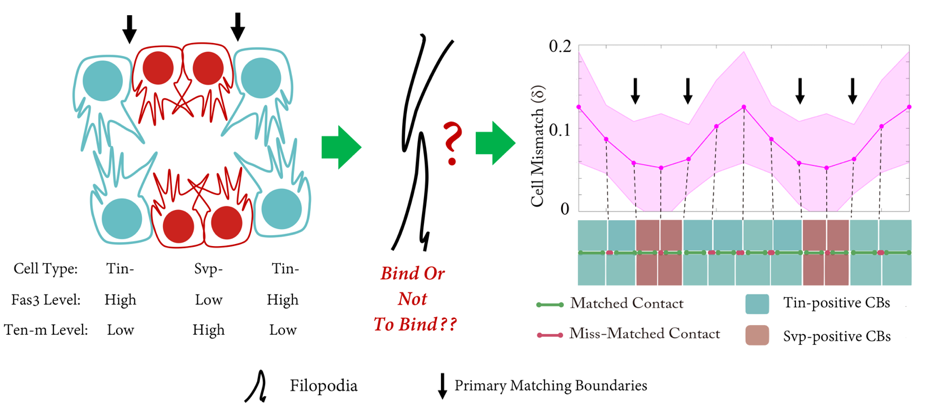
The Drosophila heart tube is constructed from two parallel lines of connected cardioblasts in the developing mesoderm, initially separated by over 100mm. The cardioblasts migrate together and create the heart tube during stage 16. The cardioblasts have distinct subtypes: Tinman-positive cells that form the heart lumen and valves; and Seven-up-positive cells that form the ostia. The Drosophila cardiogenesis ‘dating algorithm’ has Tinman and Seven-up-positive cardioblasts lined up in a repeating 4-2 pattern (Figure 1). As the contralaterally opposing lines of cardioblasts come together, they only match up with cells of the same type (Figure 1). We set out to investigate: how does this happen, and why?
Figure 1: Schematic of embryonic Drosophila heart formation. As the cardioblasts approach each other (left), their filopodia interact (middle), and eventually form precisely matched boundaries (right). Tin-: Tinman-positive cardioblasts (CBs); Svp-: Seven-up- positive cardioblasts (CBs).
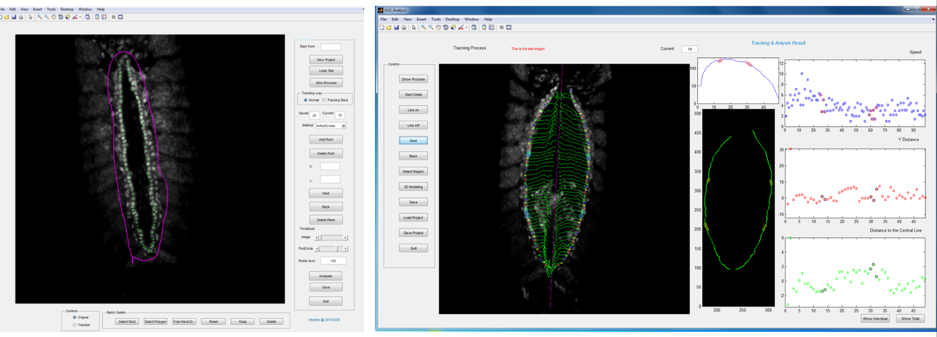
Primary ‘matchmaker’ Shaobo Zhang, as part of an undergraduate research project, had the job of figuring this out. He was working under tough conditions: less than a year old, the lab had few reagents and no fly room. Nevertheless he got his hands on a Hand::GFP line, which expresses a marker for cardioblasts in the Drosophila embryo, and developed cell tracking software to see how the cells migrate during heart formation (Figure 2). The project proved compelling enough to persuade Shaobo to join the lab (https://mbi.nus.edu.sg/timothy-saunders/) for his PhD studies, seemingly undaunted that the PI was a theoretical physicist. (It probably helped that by then we had a mini fly room – albeit with three levels of security to stop any flies escaping, Figure 3).
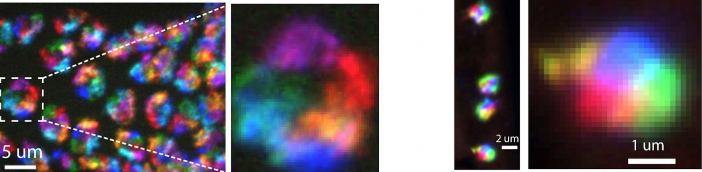
Shaobo noticed that – rather like competitors on Love Island – cardioblasts could change the partner they coupled up with quite abruptly. As the team looked more carefully, we realised that cells were sampling their local environment using filopodia protrusions. However, not all cells were equally good at finding their partners precisely. The best matched cells were at the boundary between the Tinman- and Seven Up- positive cardioblasts.
Figure 2: Cell tracking of cardioblasts with home-developed tracking software. On right, the green lines represent the cell tracks. Note that as cells approach their counterparts the tracks move more.
Figure 3: Our mini fly room (right) with three security levels to stop flies escaping (left)
But what factors helped cells decide which of the opposing cells was the right one for them? We notice that the filopodia forming strong connections were generally from the Tinman-positive cells. In contrast, the filopodia interactions between Seven Up-positive cells were distinctly unromantic, and rather more like Neymar and Ronaldo having a fight – a lot of arm-waving and drama, but no meaningful contact.
Shaobo reasoned that cell-cell adhesion molecules may be differentially expressed in cardioblasts to facilitate the selective filopodia adhesion. At this point he turned to the literature, as neurobiologists have been investigating neuronal cell matching for years and a range of known “matching” molecules are known, though the dynamic mechanisms through which they act are not fully understood.
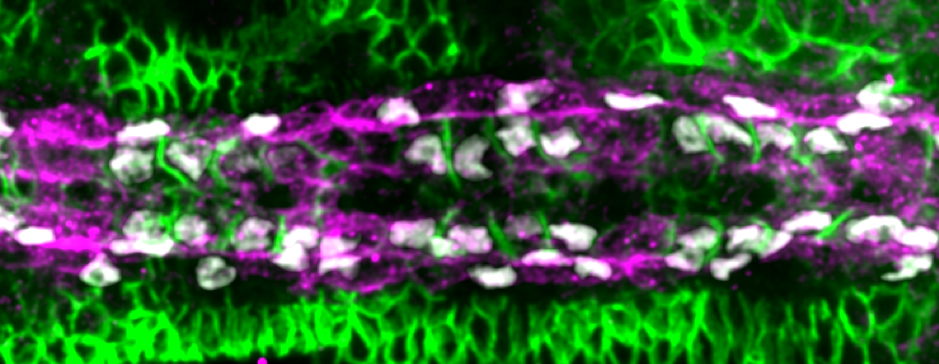
Figure 4: Antibody staining of Fas3 (green), Ten-m (magenta) and Tinman (white) in heart cells during stage 16.
Performing a mini-screen of these targets, we found that Fasciclin III (Fas3) stood out. Fas3 is a homophilic adhesion molecule (Figure 4), which was more highly expressed in the Tinman-positive cardioblasts. Perturbing the expression pattern of Fas3 resulted in perturbations of their filopodia binding activities, leading to increased cell mismatch.
At this point, it looked like we had found the molecule driving cell matching (a bit like alcohol at a student party). However, when we looked at fas3-/- mutants we noticed only a small defect in cell matching (unbelievably, dating can also occur sober). Returning to our screen results, we noticed that the adhesion molecule Ten-m (also known as Teneurin-m or Tenascin-m – sometimes naming conventions really need to be sorted out) was upregulated in Seven-up positive cells (Figure 4). After some painful crosses, Shaobo produced the double mutant of Fas3 and Ten-m, which, thankfully, had a significant matching phenotype. Therefore, it appears that heart cells use two (partially redundant) adhesion molecules to ensure they find the right partner.
But how is this differential expression pattern genetically regulated? To answer this, we turned to Dr David Garfield at Humboldt University in Berlin (https://www.garfieldlab.org/). By looking at putative enhancers specific to mesodermal and neuronal tissue, he identified potential tissue-specific enhancers for Fas3. Shaobo made reporter lines to test whether these distinct regions correspond to the specific expression patterns for Fas3. Thankfully they did, with specific expression in cardioblasts (with differential expression in distinct cell types) and neurons. So – for cardioblasts anyway – the odds of pair-bonding are partly a matter of genetic destiny.
This project was a lot of fun as we explored how both mechanical and genetic mechanisms interplayed to regulate the precise cell matching and help forming the properly structured heart. Given the conservation of many of the genes involved in early heart formation, we are hopeful that this will have relevance to vertebrate systems. More interestingly, this shows a simple but potentially general dynamic mechanisms of constructing specific cell-cell connections in biological systems. We’ll keep you posted if we crack the human relationship code too.
Reference
Zhang, S. et al. (2018) ‘Selective Filopodia Adhesion Ensures Robust Cell Matching in the Drosophila Heart’, Developmental Cell, 46(2), p. 189–203.e4. doi: 10.1016/j.devcel.2018.06.015.
 (No Ratings Yet)
(No Ratings Yet)

 (2 votes)
(2 votes)







 (4 votes)
(4 votes){kind=link}