On Growth and Form at 100: Perspectives from the field
Posted by the Node, on 28 November 2017
What explains the lasting legacy of D’Arcy Thompson’s ‘On Growth and Form’? A century on from the publication of the first issue, we reached out to authors from Development’s special issue celebrating the centenary and asked what the book meant to them.

Arkhat Abzhanov
Natural History Museum (London) & Imperial College London
“There are ideas which make their impact and give way to new and better ideas. They have their day and disappear into history. However, there also ideas that have a much more profound and enduring effect and continue to influence the way we think for many yeas, sometimes for generations. “On Growth and Form” by D’Arcy Thompson is a book full of such brilliant everlasting ideas (and so eloquently written). I remember reading it for the first time as a half-fledged undergraduate awed by the notion that living organisms and their parts do not have “random” shapes but could be related to each other in precise mathematical terms. What could be a more powerful and visual evidence for evolution than a gradual transformation of a skull of a primitive horse into a modern one via a series of shapes of the intermediate species? This book asked many important questions but gave few concrete answers, instead outlining ways of addressing problems. My own current research is very much driven by the quest to explain Thompson’s “laws of growth” and their role in evolution while using his “theory of transformations” in the form of modern geometric morphometrics. For example, the origin and subsequent diversification of the uniquely-shaped skull of modern birds can be better understood by comparing avian skull shapes with each other and with their reptilian ancestors – all within the same “morphospace”, the concept invented by D’Arcy Thompson. Thus, his ideas in many ways are still surprisingly relevant and stimulating today as ever before. I hope that the 100th anniversary will bring this book more attention from the new generation of scientists curious about biological shapes.”
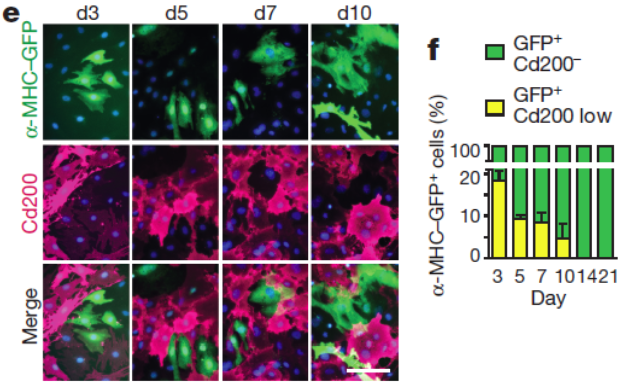
Arkhat’s review – The old and new faces of morphology: the legacy of D’Arcy Thompson’s ‘theory of transformations’ and ‘laws of growth’
Douglas DeSimone
University of Virginia
“I spent several summers in the early 1980’s undertaking my doctoral research on early sea urchin development at the Marine Biological Laboratory (MBL) in Woods Hole. I remember fondly the diverse array of sea life that John Valois and his crew in the supply department would collect for seasonal investigators over the course of each summer. Their labors afforded opportunities to observe and handle not only adult marine animals – many of which I had never seen alive before – but also, on occasion, their eggs and embryos. It was against this backdrop that I developed a career-long fascination with the problem of morphogenesis, and first became acquainted with D’Arcy Thompson’s “On Growth and Form”. Back then, there was a small bookstore on Water Street that was unusual for carrying not only the light summer reading demanded by island-bound tourists, but also a selection of titles reflective of the greater MBL and Woods Hole Oceanographic Institute scientific communities. One August, as the exodus of summer investigators, students and tourists began to transform Woods Hole back to its less frenetic state, I purchased a paperback edition of Thompson’s book, which I began reading on the beach and would continue back at my home institution that Fall. Thompson’s perspective inspired me to appreciate the problem of development as more than an unfolding of genetically-encoded programs but one that is also subject to physical rules and open to mathematical description. In many ways, I feel that my research and that of others interested in morphogenesis has moved inexorably toward a view of embryos as biomechanical “machines”. Thompson likely could not have imagined the modern tools and approaches now being developed to measure, analyze and model forces generated by cells and tissues but it is safe to say he would have approved of where the field has headed since the publication of his influential work.”
Paper from the DeSimone lab – ‘Mechanical and signaling roles for keratin intermediate filaments in the assembly and morphogenesis of Xenopus mesendoderm tissue at gastrulation‘
François Graner & Daniel Riveline
Université Denis Diderot, Paris and IGBMC, Strasbourg
“DR is an experimentalist biophysicist, working on acto-myosin as well as cell adhesion and motility in vitro; FG is at the interface between tissues and foams, and between theory and experiments. We have been knowing each other 20 years ago, while we were in the same lab in Grenoble, and our first discussions in 1999 made already clear that MDCK cells monolayers cellular outlines were not following rules of shapes for bubbles (see our original MDCK image below).
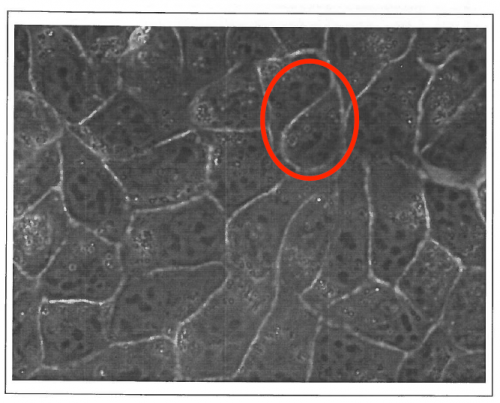
We both then moved to different scientific sites, and we started to interact again in 2010 around a new Master program launched by DR in Strasbourg University, where we gave interdisciplinary lectures to the same students. DR suggested to share classics pages with students on considering living matter as a matter, such as Thompson book proposes, with modern echoes in developmental biology. FG was reluctant in considering these pages as actually seminal: he rather considered that they had been causing a waste of time over the century, by triggering inadequate searches for simplicity and perfection. These distinct appreciations of ‘On Growth and Form’ nurtured between us an animated debate. We decided to focus on two chapters which were central for us, i.e. on shape and packing of cells in tissues. Our disagreement was so strong that it was difficult to imagine that we would agree on a common flow; initially, we had no agenda whatsoever to wrap up this discussion in an article one day. We turned this debate into a scientific approach, by carefully listing specific arguments and counter-arguments.
And then, we both moved from our original respective postures. DR recognized that Thompson had been merging phenomena under the same umbrella, with often analogy as a guideline and limited care about physical mechanisms. FG acknowledged the influence of Thompson in general and even on his own work for several aspects, and particularly the illuminating ability of Thompson to act as honest experimentalist representing tissues through their essential read-outs – cell contours, representing them as they actually are. We had just decided to write an article when, by coincidence, Development launched a call for its special issue.
A long way has been done since our very first discussions, and we view a posteriori our debates as one of the most productive discussions we had in recent years, with no other ambition than understanding the text and its scope a century after its first publication. And we can only encourage others to perform the same patient task for other chapters of this book or other classical texts in Biology.”
François and Daniel’s review – ‘The Forms of Tissues, or Cell-aggregates’: D’Arcy Thompson’s influence and its limits‘
Olivier Hamant
University of Lyon
“The field of development has reinvented itself several times over the past decades, building from grafting/morphogen-centered experiments to molecular genetics, cell biology and now including more quantitative and predictive aspects, through biophysics and computational modeling. With the rise of quantitative approaches, the current era is very much in line with D’Arcy Thompson persistent words:
“Dreams apart, numerical precision is the very soul of science, and its attainment affords the best, perhaps, the only criterion of the truth of theories and the correctness of experiments”
D’Arcy Thompson certainly was a (maybe too) strong advocate of the role of physical forces in development, but as we are collecting more and more evidence of a widespread role of mechanical signals in morphogenesis and cell behavior, his take on development also offers a source of inspiration for the young generation of interdisciplinarians in development.
One key quote that was influential to me is a simple one:
“The form of an object is a “diagram of forces””
The simplicity of these words has many implications. It underlines the fact that changing shape is not only a geometrical problem: it also involves a structural change, and thus, that changing patterns of forces must be managed by cells. Not as immediately obvious, this statement also implies that a living object will usually try to resist such forces. There are now many examples of cells displaying such resistance strategies, from changes in cell growth rate, to cell polarity or extracellular matrix stiffening. This also is true at tissue and organ scales, the most obvious example being Wolff’s law, which is mentioned in D’Arcy’s book, where bones build their intricate trabeculae network along maximal mechanical stress lines. We, and others, found similar responses in plants, with cells and tissues reinforcing their structure in the direction of maximal stress too, making this statement from physics, a true biological theorem. D’Arcy Thompson was a supporter of the organismal theory of development, and this quote is also consistent with that option, diagrams of forces spreading, and arguably, synchronizing, populations of cells. While this leaves out many subtleties, it remains a very powerful reminder of the fact that cells manage communication, nutrition, reproduction, etc. and while doing all this, also need to maintain their own structural integrity, which calls for synergies between biochemical and mechanical signalling.
One last take home message from D’Arcy Thompson’s book is his didactic choice: the use of many analogies makes his book very accessible to a wide audience, truly bridging biology, mathematics and physics. This is a remarkable piece of work, and despite several questionable interpretations 100 years down the road, “On growth and form” remains a landmark in developmental biology, and very much worth a read today, for biologists, mathematicians and physicists alike.”
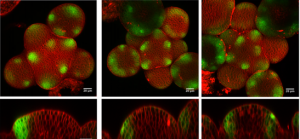
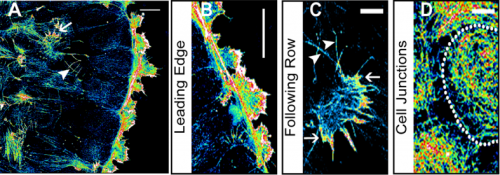
Paper from the Hamant lab – ‘Phyllotactic regularity requires the Paf1 complex in Arabidopsis‘
Akatsuki Kimura & Kazunori Yamamoto
SOKENDAI & National Institute of Genetics, Mishima, Japan
AK “When I was a post-doc, I first read “On Growth and Form” in the Japanese version translated by Dr. Tomomichi Yanagida et al. and published in 1973. In the translators’ preface, Dr. Yanagida wrote the following: “Initially, we were not sure whether the book is worth translating, as the book is about 70-years old and the biology had been progressed dramatically since then.” For me, having read many textbooks on molecular biology, reading this book was an eye-opening experience. Explaining features in sizes and shapes of various organisms at the cellular level (or beyond), using mathematics and physics was an area I wished to pursue as a scientist. Many topics of study in this 100-year-old book are currently pursued by many quantitative biologists. In fact, our manuscript in this special issue of Development was inspired from the chapter “The Forms of Tissues, or Cell-aggregates.” I hope that our manuscript has provided a novel insight into the topic covered in this chapter. Dr. Yanagida concluded the preface as follows: “Nowadays, there is a flood of biology textbooks, but this book is one of a few books that is truly inspirational”; I completely agree with this statement.”
KY “I learned about this book for the first time when I was a graduate student. At that time, I was skeptical of pursuing research in molecular biology because I found it difficult to believe that the essence of the organisms I wished to work on could be revealed by simply focusing on the function of a single gene. The book looked at the question from a different point of view. Therefore, I found the philosophy of the book attractive. Indeed, for many people who like to watch organisms including myself, one of the interesting aspects of organisms is the various beautiful structures they create. The premise of the book that the shape of different structures, such as a cluster of cells, the skeleton of planktons and sponges in the ocean, and the body shape of jellyfish, might be explained by surface properties of liquids, is indeed enthralling. The association of surface tension with the broad range of biological shapes, from cell shapes (micron-size) to animal body shapes (meter-size), intrigues me. Now, I deeply respect D’Arcy Thompson’s abundant knowledge and speculations regarding organismal morphology based on physical and mathematical viewpoints, and I shall remember him every time I observe dynamic behaviors of organisms under a microscope.”
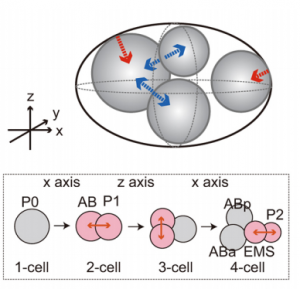
Kazunori and Akatsuki’s paper – ‘An asymmetric attraction model for the diversity and robustness of cell arrangement in nematodes‘
Rusty Lansford
Keck School of Medicine, University of Southern California & Children’s Hospital Los Angeles
“I first ‘read’ Thompson’s ‘On Growth and Form’ as an undergrad. I found it rather daunting in a kind of War and Peace thick science text way. I tepidly thumbed through the pages, mostly stopping at interesting figures, especially the ones about spirals in nature and the accompanying mathematics. Soon I was up the N. California coast abalone diving with friends. We were there to collect and enjoy the yummy gastropods with local spirits. We always enjoyed the iridescent nacre that lines the inner shell. However, on this trip, my mind drifted back to Thompson’s figures of Haliotis, and its precise patterns of growth. My perspectives had forever changed, even on road trips with friends, to see patterns and morphometrics everywhere.”
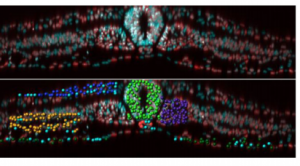
Paper from Rusty Lansford and colleagues – ‘Multi-scale quantification of tissue behavior during amniote embryo axis elongation‘
Julien Vermot
IGBMC, Strasbourg
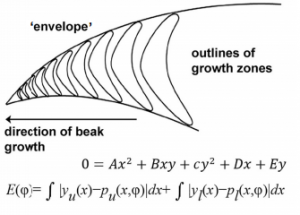
“I was phD student the first time I heard from on Growth and Form. Thomas Lecuit gave a very nice seminar in our institute. I remember him showing the famous drawings describing how environmental forces may affect the overall shape of an adult fish (it should correspond to the Figure 517 to 524). As a PhD in developmental biology, it had a strong impact on me. At the time, we were struggling with in situ hybridization and our understanding of how shape is emerging out of these expression profiles was clearly not satisfactory. I cannot say if it was the drawings themselves or if the field had already shifted, but the idea of forces affecting shape led to lots of discussion in the institute. It definitely influenced my post doctoral work and current research.”
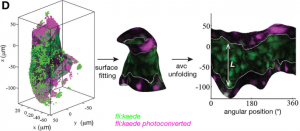
Paper from the Vermot lab – “Anisotropic shear stress patterns predict the orientation of convergent tissue movements in the embryonic heart“
Thanh Thi Kim Vuong-Brender
Labouesse Lab, IBPS, Paris
“Throughout the book, which I read in a French translation of the abridged John Tyler Bonner version, D’Arcy Thompson tried to use physics and mathematics to explain the shape of life forms. This was an audacious project, since the living world remained a big mystery at that time. D’Arcy Thompson foresaw the great utility of physics and mathematics in studying growth and form, while admitted that they could not explain all the living phenomena. He stressed the importance of material and mechanical nature of life, considering that a living form was nothing else than “a force diagram”. D’Arcy Thompson illustrated the beauty of a wide variety of species through mathematical description. He pointed out the similarities between seemingly unrelated things, linking the natural phenomena with physical laws. Modern research on morphogenesis has proved that many of these intuitions were correct.
As a researcher working on a particular morphogenetic problem (elongation of C. elegans embryo), I also look at the biological phenomenon from a physical point of view, in terms of “materials”, “forces” and “force distribution and evolution”. Like D’Arcy Thompson, I also try constantly to relate the morphogenetic mechanisms with physical laws and physical phenomena that we see in daily life. Some paragraphs of the book are directly related to my work, for example the one stating that the formation of shapes different from a perfect sphere may results from either a difference in internal pressure (inside the object) or the anisotropy of the surface envelope. The relevance to my particular case on the anisotropic deformation C. elegans embryos shows that D’Arcy Thompson’s observations can still inspire future generations of scientists. Although many of the theories in “On Growth and Form” were simplistic and unproven, the observations and reasoning are a good incentive to further investigate the nature of the phenomenon. The book deserves to be read broadly, not only for its pioneer ideas on morphogenesis but also for its enjoyable elegant writing style.”
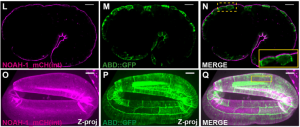
Thanh’s paper – ‘The apical ECM preserves embryonic integrity and distributes mechanical stress during morphogenesis‘
What does On Growth and Form mean to you? We’d love to hear your thoughts in the comments section below
On Growth and Form at 100 on the Node:
An interview with Matthew Jarron
Morphogenesis one century after On Growth and Form
 (3 votes)
(3 votes)




 (1 votes)
(1 votes)