Over the past year and a half, I had the opportunity to participate in the UW-Madison Kohler Fellowship Program. In this program, an artist is paired with a scientist to create a science-art fusion project, some goals of which include fostering cross disciplinary communication and increasing the accessibility of scientific concepts through art. Though I was accepted into this program as a science fellow, my partner, Sharon Tang, and I both consider ourselves artist-scientists. Sharon is a PhD candidate in the Cell and Molecular Biology program and an avid muralist, while I’m a Genetics PhD candidate and composer/musician. Together we leveraged our scientific knowledge and artistic expertise to create “Genetic Symphonies: the building Hox of life”.
“Podium”: The control panel for “Genetic Symphonies: Building Hox of life” contains 13 black buttons arranged on a music staff that was laser cut into wood. The stem of each music note is color coded to match its corresponding Hox box. Buttons must be pressed in the correct order to activate Hox gene expression in the corresponding box.
As most developmental biologists know, Hox genes are transcription factors critical for patterning the skeletal axes. They also remain one of science’s biggest mysteries; we still have yet to determine what Hox genes actually regulate to control developmental and adult homeostatic processes. In addition to the mysterious mechanisms of action, Hox genes possess unique properties (clustered chromosomal arrangement, spatio-temporal expression pattern, regional restriction) that lend nicely to artistic manipulation and exploration.
Sharon and I took advantage of these features to generate an interactive, multi-modal exhibit. In our exhibit, 13 paralogous groups of Hox genes are represented by 13 painted building blocks, affectionately referred to as “Hox boxes”. Likewise, there are 13 color-coded, randomized, buttons that control each “Hox box” on an accompanying podium. Participants must activate gene expression, via a button press, and determine the correct order of Hox gene activation (head to tail). Each correct button press produces light in the corresponding box and sounds a unique measure of music. With each successive, correct, button press participants develop their own genetic symphony and an abstract human figure via lights. By conveying development through both light and sound, we also increase the accessibility of our exhibit.
Want to know more about Genetic Symphonies: Building Hox of life? Check out Behind Building: Hox ! At this site, you can find more information about the construction, fabrication, composition, and electronics of the exhibit.
“Lights on”: When all 13 buttons are pressed in the correct order, a hidden abstract human figure illuminates from within the boxes. (3 votes) Loading...
Hydractinia symbiolongicarpus is an emerging model to understand stem cell evolution
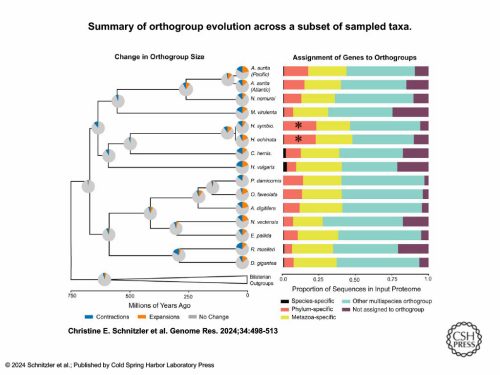
Stem cells can’t hide what they are. At least, that’s the takeaway from the newly sequenced genomes of two colonial hydroids, Hydractiniasymbiolongicarpus and Hydractinia echinata. I recently sat down with Dr. Christine Schnitzler of UF’s Whitney Laboratory for Marine Bioscience to talk about her experience assembling these genomes — a huge collaboration that started pre-pandemic, slowed down during the pandemic, and finally culminated in a paper published in Genome Research in March. Our conversation covered topics spanning from the logistics of large collaborations (including the need to find the right people), the shared molecular vocabulary that stems from genome projects, and the scientific merit of studying emerging systems.
Hydractina are colonial cnidarians, a group that consists of anthozoans (sea anemones and corals), schyphozoans (co-called “true” jellyfish), cubozoans (box jellyfish), and hydrozoans (Hydra and Hydractinia). Cnidarians have an informative phylogenetic position as sister to bilaterians. In addition to providing evolutionary insights, many cnidarian species — including Hydractinia symbiolongicarpus — are exceptionally good at regenerating. Schnitzler’s lab is studying Hydractinia symbiolongicarpus to understand how interstitial stem cells, or i-cells, impact an organism’s ability to regenerate.
“All genome projects are huge,” said Schnitzler. “I would never have been able to do this myself.”
This particular project has been 10 years in the making. In that time, sequencing technologies and analyses kept changing. The research team opted to swap the Illumina short-read platform for Pac-Bio long-read sequencing to dig deeper into two unconventional model organisms: Hydractinia echinata and Hydractinia symbiolongicarpus.
The genomes of these two cnidarians had a few surprises. First, the two genomes were quite different in size — Hydractinia echinata was 775MB and Hydractinia symbiolongicarpus was 514MB. And that’s not just based on sequence. The research team isolated single cells, stained nuclei with propidium iodide and ran them through a flow cytometer against a known standard. Second, based on the researchers’ analysis, these two species’ genomes diverged around 19 million years ago. That may seem like a long time, but if you consider that two different strains of the same species of jellyfish are estimated to have diverged 45 million years ago, then a true species divergence 19 million years ago is surprisingly recent.
“I thought that was pretty cool,” said Schnitzler.
Genomes offer a common language for biologists to understand similarities (and, importantly, differences) between species. To that end, Schnitzler’s team took a lot of time and care into making this resource available for anybody with even a cursory interest. They also developed a web portal for both Hydractinia species that allows curious biologists to dive deeper into a very, very granular level of gene evolution between two closely related genomes. They also generated a single cell RNAseq browser.
“That’s kind of fun. The whole point is to make this genome accessible and useful to the community,” said Schnitzler. “It was a priority for us to build that resource, which was not easy,” she added with a laugh.
“Collaboration is key. And having the right people is very helpful.”
At the end of our discussion, I gave Schnitzler a chance to respond to the “Questions For The Author” from PreLighter Isabella Cisneros who spotlighted the preprint last September (check out the PreLight article here). I’ve included her questions, numbered 1–3, as well as Schnitzler’s responses below.
Questions For The Authors (from PreLighter Isabella Cisneros):
1. How do you reconcile the large number of shared i-cell marker genes with the higher proportions of phylum-specific and cnidarian-specific genes in the H. symbiolongicarpus genome?
We took the entire genome and ran an orthology clustering of all predicted proteins from 49 other animals, including 16 cnidarians. We put them into bins once we got the clusters back. Are these genes in a “multi-species orthogroup?” That means they group with genes from animals outside of cnidarians, so they’re shared more widely. A lot of the other categories were cnidarian-specific.
This graph shows just the cnidarians and a few other animal outgroups. Those two [asterisked] red bars are further out than all the other cnidarians — they had the highest proportion of genes in their genome that were specific to their phylum.
If you apply that same clustering to the most highly expressed genes that are specific to just the i-cells, we found that i-cell genes are mostly shared with other animals (that was the title of the paper).
How do I reconcile that? I think it means that stem cells are exactly what they are.
They’re undifferentiated cells that have to use basic cell characteristics — cell cycle genes, genes that help with proliferation, genes that are involved in maintaining stemness — and these are universal things that all animals have developed and retained throughout evolution.
That does not mean that stem cells of Hydractinia are exactly homologous to the stem cells of other animals. Cell types can evolve very quickly, but the underlying genes that make stem cells stem cells are very highly conserved.
There are a lot of cnidarian-specific genes in their genomes. It’s just in their stem cells, they’re not using those very much. But when you think about stem cells, they’re not really unique to cnidarians. All animals have some type of a stem cell — that’s just how it works.
2. Towards the end of the preprint, you claim that it remains unclear whether other animals share the same toolkit of genes, or whether these toolkits are instead partially overlapping. While further studies will be necessary to determine this, at this stage, what do you anticipate to be the case?
It’s very hard to relate cell types and say they have a shared evolutionary origin of the cell type. If you look at stem cells in Hydractinia and then look at stem cells in planarians, they have a lot of shared characteristics. But what about all the animals in between? Where did their stem cells go? I think that’s a very interesting question.
I think getting down to this core level of genes and gene regulatory networks that are controlling these types of cells, to me, might be more interesting and more informative from an evolutionary perspective than trying to absolutely say this cell type first arose at some point and then shifted. It’s not really about the cell types. It’s more about the core genes that are involved and finding out their function. And it’s really hard. I think the only way really is to drill down on function within several animals and then try to see if they relate to each other.
It’s good to look at informative positions on the tree, as a lot of people try to do, and try to gain as much insight as you can from looking at those different places. I don’t think there’s going to be 100% overlap. But I think there will be themes that emerge — categories of genes that may be similar. You could group them by orthology, but maybe not exactly by BLAST.
It’s a question for the future. I think people are trying to tackle it, but it’s going to take more than these genome-wide approaches. We’ll have to go back to the lab and do functional testing to get at those questions.
3. Given the different evolutionary trajectories that H. symbiolongicarpus and H. echinata have followed since divergence, what kinds of studies would be better suited for each species, if any? What could be gained by using both species in a comparative framework?
Most people have dropped H. echinata as a research organism. It’s got a bigger genome that’s not as well assembled. And no one’s maintaining them in the lab, anymore. They’re just harder to keep in culture long term. There may be some ecological and other questions that might be more interesting with echinata, and it would be cool if someone picks it up now that the genome is available.
On the other hand, H. symbiolongicarpus just grows and grows and grows indefinitely. After years and years of being in culture, we can still get them to spawn on a weekly basis and get tons of embryos. It also has a smaller genome that ended up with a better assembly.
Smaller genome, easier lab culture — all of that screams, “Work on this one!”
So for us, the path forward is symbiolongicarpus. Now we can talk and really understand each other when we’re talking about a particular gene or a particular process. Now we have this resource. It’s a starting point.
One interesting thing to think about is what is unique about Hydractinia biology? I like Hydractinia because it is different. It’s colonial with a polymorphism of polyp types. We have feeding polyps, sexual polyps, defensive polyps. Because we have this diversity just within a single colony, there are a lot more questions to ask. It’s a different type of development, a different kind of asexual reproduction.
The other thing that I think is super cool is its lifecycle. It’s a hydrozoan that has lost the medusa stage. So there’s no jellyfish stage. Its next closest sister group is Podocoryna carnea, which does have a polyp and a medusa stage. That genome is being sequenced now. So when we have these two genomes — Podocoryna and Hydractinia — you can now start to understand the difference between a hydrozoan genome that produces the medusa jellyfish and the genome that doesn’t. I think some of these comparative studies with new genomes are really, really exciting. And I think with unlimited resources, we would have perfectly assembled genomes for those two groups and start doing more experiments to try to understand the unique biology of Hydractinia.
One thing our paper does not really focus on is regeneration. Hydractinia symbiolongicarpus is a model of regeneration. There’s a huge amount of knowledge we can learn by studying regeneration in this animal. There’s been some seminal ground laying papers about regeneration in this model, but doing updated studies, which we have some data, hopefully soon we can update and talk more about how this animal achieves its amazing regenerative abilities.
At the speakers’ discretion, the webinar will be recorded for viewing on demand. To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
In this SciArt profile, we meet Friedrich Bliem, who has a background in cell biology and scientific illustration and has been creating “Art in Science” paintings for decades.
Can you tell us about your background and what you work on now?
I am Austrian and have spent a total of 24 years in the USA, UK and Australia, where I was raised. As the son of a freelance artist, I practiced the skills of drawing and painting from an early age and in my teens focussed on later enrolling in the University of Applied Arts, Vienna. Music had also been a passion and I co-founded the successful Austrian music group, Misthaufen, which is still active today.
Despite my interests in both music and art, I also had an innocent fascination for science based on the link between art and music. So, in pursuit of this interest, I chose first to enroll in a natural sciences and engineering course at a Viennese university, which was then renamed Biotechnology.
Concomittantly, I continued my art training with non-formal education, especially under the expressionist artist, Rudolf Macek.
After completing my university studies in 1981, I moved to Cambridge, England, where I continued to paint.
I had always had a passion for drawing cartoons as it allowed me to reflect on my social environment without having to say sorry. This passion motivated me to approach the Elsevier Publications office in Cambridge with some ideas and examples of my work. Soon I found a successful niche as scientific illustrator and cartoonist for various publishers.
Encouraged by the Cambridge environment, as well as my work as illustrator, I began working on scientific, especially cell biological, subject matters. This culminated in a solo exhibition in Cambridge, UK, 1984 (“Art in vivo”). As such, I might be said to be a fore-runner of today´s so-called SciArt movement.
However, in 1988 I decided to devote my time to science, before again returning to art. This period lasted 25 years!
Cells Tissues Organs Watercolour, 30×45 cm 1987
Were you always going to be a scientist?
No, not really. Like so many youngsters I had a chemistry set and developed a fascination for chemistry, but the continuous smell of turps and oil in our house always drew me back to drawing and painting. Music was simply another form of expression. The intensive engagement in music led me to question the connection between music and art. I was convinced there was a physical connection, but also realised that I simply didn´t have the theoretical knowledge to proceed. This was the sole reason for choosing a study programme with a broad curriculum both in biology and engineering over art school.
Evolution Oils on panel, 50×70 cm 2023
And what about art – have you always enjoyed it?
Well, yes and no. I wouldn´t speak of joy. My relationship with Art was set in a much deeper fashion as just to be enjoyed. It was an intuitive means of expression that came easily, be it fine art or music, which found some success, even in school days, and of course in which I reveled. But it also stemmed from an admiration for my father and our complex relationship.
Fungal scene Watercolour, 30×40 cm 1988
What or who are your most important artistic influences?
Of course my father, whose work was largely impressionistic. And Rudolf Macek, who was clearly an expressionist of the 1950´s and 60´s. And from the public figures first and foremost they were and are Franz Marc, Wassily Kandinsky, August Renoir; painters of the romantic period, such as Ferdinand Waldmüller, and of course the old masters such as Jan Vermeer or Salomon van Ruysdael.
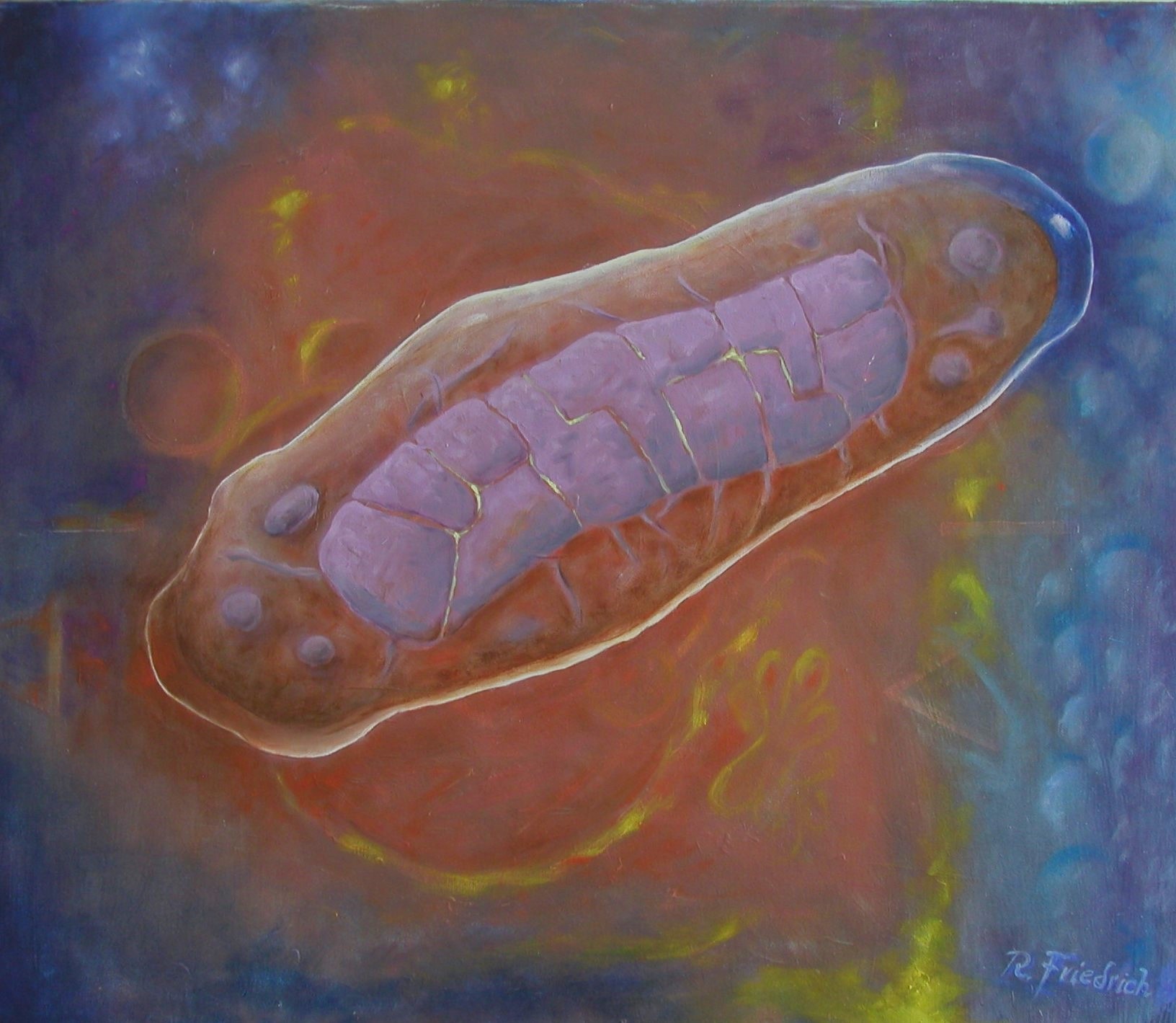
Mitochondrium 4 Oils on linen, 50×60 cm 2024
How do you make your art?
Today I only paint, on canvas or panels, in oils, sometimes with a draft drawing in acrylic. The Cambridge atmosphere lends itself to watercolours, which is well accepted.
Neuron Acrylics and oils on panel, 70×135 cm 1985
Does your art influence your science at all, or are they separate worlds?
Whereas my scientific subjects definitely reflect my knowledge of cell biology, art has tuned my visual sensitivity to aesthetics in science and engineering, e.g. a factory with its piping systems or SEM photos of fungal hyphae and conidia. This has even, to some extent, determined significant professional choices I have made.
Compostion of the Self Oils on panel, 50×60 cm 2024
What are you thinking of working on next?
I will continue to expand on my scientific themes, one of which is the concept of evolution. But I also have a “macrobiological” subject, in which I seek to capture the essence of animals in their environment.
On the topic of plant development and chaired by Development Editor, Dominique Bergmann (Stanford University).
Wednesday 31 July – 16:00 BST
Martina Cerise (Max Planck Institute for Plant Breeding Research) ‘In plants, the organization of the apical stem-cell niche changes dynamically during the floral transition’
Vicky Spencer (Gregor Mendel Institute) ‘‘How to build a plant: Small meristems have big consequences’
Gwendolyn K. Kirschner (The James Hutton Institute) ‘Control of the root gravitropic set-point angle in barley’
At the speakers’ discretion, the webinar will be recorded for viewing on demand. To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
Bechara Saykali, Andy D. Tran, James A. Cornwell, Matthew A. Caldwell, Paniz Rezvan Sangsari, Nicole Y. Morgan, Michael J. Kruhlak, Steven D. Cappell, Sergio Ruiz
Lieke Golbach, Tanumoy Saha, Maria Odenthal-Schnittler, Jenny Lücking, Ana Velic, Emir Bora Akmeric, Dorothee Bornhorst, Oliver Popp, Philipp Mertins, Felix Gunawan, Holger Gerhardt, Boris Macek, Britta Trappmann, Hans J. Schnittler, Milos Galic, Maja Matis
Anastasia Chugunova, Hannah Keresztes, Roksolana Kobylinska, Maria Novatchkova, Thomas Lendl, Marcus Strobl, Michael Schutzbier, Gerhard Dürnberger, Richard Imre, Elisabeth Roitinger, Pawel Pasierbek, Alberto Moreno Cencerrado, Marlene Brandstetter, Thomas Köcher, Benedikt Agerer, Jakob-Wendelin Genger, Andreas Bergthaler, Andrea Pauli
Andrew Katznelson, Blake Hernandez, Holly Fahning, Jingchao Zhang, Adam Burton, Maria-Elena Torres-Padilla, Nicolas Plachta, Kenneth S. Zaret, Ryan L. McCarthy
Jordy Dekker, Wendy Lam, Herma C. van der Linde, Floris Ophorst, Charlotte de Konink, Rachel Schot, Gert-Jan Kremers, Leslie E. Sanderson, Woutje M. Berdowski, Geeske M. van Woerden, Grazia M.S. Mancini, Tjakko J. van Ham
Mary-Bronwen L. Chalkley, Lindsey N. Guerin, Tenhir Iyer, Samantha Mallahan, Sydney Nelson, Mustafa Sahin, Emily Hodges, Kevin C. Ess, Rebecca A. Ihrie
Archana Kamalakar, Brendan Tobin, Sundus Kaimari, M. Hope Robinson, Afra I. Toma, Timothy Cha, Samir Chihab, Irica Moriarity, Surabhi Gautam, Pallavi Bhattaram, Shelly Abramowicz, Hicham Drissi, Andrés J. García, Levi B. Wood, Steven L. Goudy
Jessica Honorato Ribeiro, Emre Etlioglu, Jasmine Buset, Ann Janssen, Hanne Puype, Lisa Berden, André Claude Mbouombouo Mfossa, Winnok H. De Vos, Vanessa Vermeirssen, Sarah Baatout, Nicholas Rajan, Roel Quintens
C Parikh, RA Glenn, Y Shi, K Chatterjee, EE Swanzey, S Singer, SC Do, Y Zhan, Y Furuta, M Tahiliani, E Apostolou, A Polyzos, R Koche, JG Mezey, T Vierbuchen, M Stadtfeld
Ying Xu, András Székely, Steffen Ostendorp, Saurabh Gupta, Melissa Tomkins, Lei Yang, Federico Apelt, Yan Zhao, Eleni Mavrothalassiti, Linda Wansing, Julia Kehr, Eleftheria Saplaoura, Friedrich Kragler
Javier Cabrera, Alvaro Sanchez-Corrionero, Angels de Luis Balaguer, Laura Serrano-Ron, Cristina del Barrio, Pilar Cubas, Pablo Perez-Garcia, Rosangela Sozzani, Miguel Moreno-Risueno
Martin William Battle, Scott Fraser Ewing, Cathryn Dickson, Joseph Obaje, Kristen N. Edgeworth, Rebecca Bindbeutel, Rea Antoniou Kourounioti, Dmitri A. Nusinow, Matthew Alan Jones
Maryam Rahmati Ishka, Hayley Sussman, Jiantao Zhao, Eric Craft, Li’ang Yu, Andrew Nelson, Miguel Pineros, Mark Tester, Dorota Kawa, Zhangjun Fei, Magdalena M. Julkowska
Yolanda Durán-Medina, David Díaz-Ramírez, Humberto Herrera-Ubaldo, Maurizio Di Marzo, Andrea Gómez Felipe, J. Erik Cruz-Valderrama, Carlos A. Vázquez, Herenia Guerrero-Largo, Lucia Colombo, Ondrej Novak, Stefan de Folter, Nayelli Marsch-Martínez
J.-F. Trontin, M.D. Sow, A. Delaunay, I. Modesto, C. Teyssier, I. Reymond, F. Canlet, N. Boizot, C. Le Metté, A. Gibert, C. Chaparro, C. Daviaud, J. Tost, C. Miguel, M.-A. Lelu-Walter, S. Maury
Jaclyn A. Adaskaveg, Chaehee Lee, Yiduo Wei, Fangyi Wang, Filipa S. Grilo, Saskia D. Mesquida-Pesci, Matthew Davis, Selina C. Wang, Giulia Marino, Louise Ferguson, Patrick J Brown, Georgia Drakakaki, Adela Mena-Morales, Annalisa Marchese, Antonio Giovino, Esaú Martínez, Francesco Paolo Marra, Lourdes Marchante Cuevas, Luigi Cattivelli, Paolo Bagnaresi, Pablo Carbonell-Bejerano, Grey Monroe, Barbara Blanco-Ulate
Ling S. Loh, Kyle A. DeMarr, Martina Tsimba, Christa Heryanto, Alejandro Berrio, Nipam H. Patel, Arnaud Martin, W. Owen McMillan, Gregory A. Wray, Joseph J. Hanly
Hui Xiao, Adam Stevens, Helen L. Smith, Karolina Szczesna, Maria Keramari, Gregory Horne, Andras Dinnyes, Susan J. Kimber, Pietro Lio, Daniel R. Brison
Rachel A. Glenn, Stephanie C. Do, Karthik Guruvayurappan, Emily K. Corrigan, Laura Santini, Daniel Medina-Cano, Sarah Singer, Hyein Cho, Jing Liu, Karl Broman, Anne Czechanski, Laura Reinholdt, Richard Koche, Yasuhide Furuta, Meik Kunz, Thomas Vierbuchen
Ary Marsee, Arabela Ritchie, Adam Myszczyszyn, Shicheng Ye, Jung-Chin Chang, Arif Ibrahim Ardisasmita, Indi P Joore, Jose Castro-Alpízar, Sabine A Fuchs, Kerstin Schneeberger, Bart Spee
Samantha Lapehn, Sidharth Nair, Evan J Firsick, James MacDonald, Ciara Thoreson, James A Litch, Nicole R Bush, Leena Kadam, Sylvie Girard, Leslie Myatt, Bhagwat Prasad, Sheela Sathyanarayana, Alison G Paquette
The LAG-R framework (Laboratory Animal Genetic Reporting) has just been published in Nature Communications with Lydia Teboul, head of our Molecular Cell Biology team at the Mary Lyon Centre at MRC Harwell and Guillaume Pavlovic, Head of Unit, Genetic Engineering and Model Validation Department at the Institut Clinique de la Souris– PHENOMIN– IGBMC, as corresponding authors. The LAG-R framework is a set of guidelines to support more complete documentation of the genetic make-up of animals that are used in research, with the aim of bolstering reproducibility, reliability, and overall scientific rigour.
The biomedical research community is addressing many different factors that lead to problems with reproducibility, including via the implementation of the PREPARE guidelines, which aim to improve experimental design, and the ARRIVE guidelines, which aim to improve reporting of animal research experiments. However, a need remains for a more comprehensive description of the genetics of research animals, as differences in genetic background that are too often perceived as subtle can have a significant impact on phenotype and genetic modifications are rarely fully documented.
The LAG-R guidelines are designed to improve the documentation that is associated with animal research and to be applicable to the full range of animal species used. Standardising and improving genetic documentation will enhance research reliability and reduce wastage of resources and animals by cutting down on the reconstitution of missing information or on follow-up experiments that unknowingly use animals with different genetics.
The authors said: “The LAG-R Guidelines are not about influencing what models researchers use for their research. They are about standardising the way the genetics of these animals are documented. This framework is intended to be simple to adopt and takes into account the diversity of research environments where animals are used. Documentation is a key part of research reproducibility!“
The authors are working to create a publicly available resource web page to facilitate the uptake of the guidelines by the community. Watch this space!
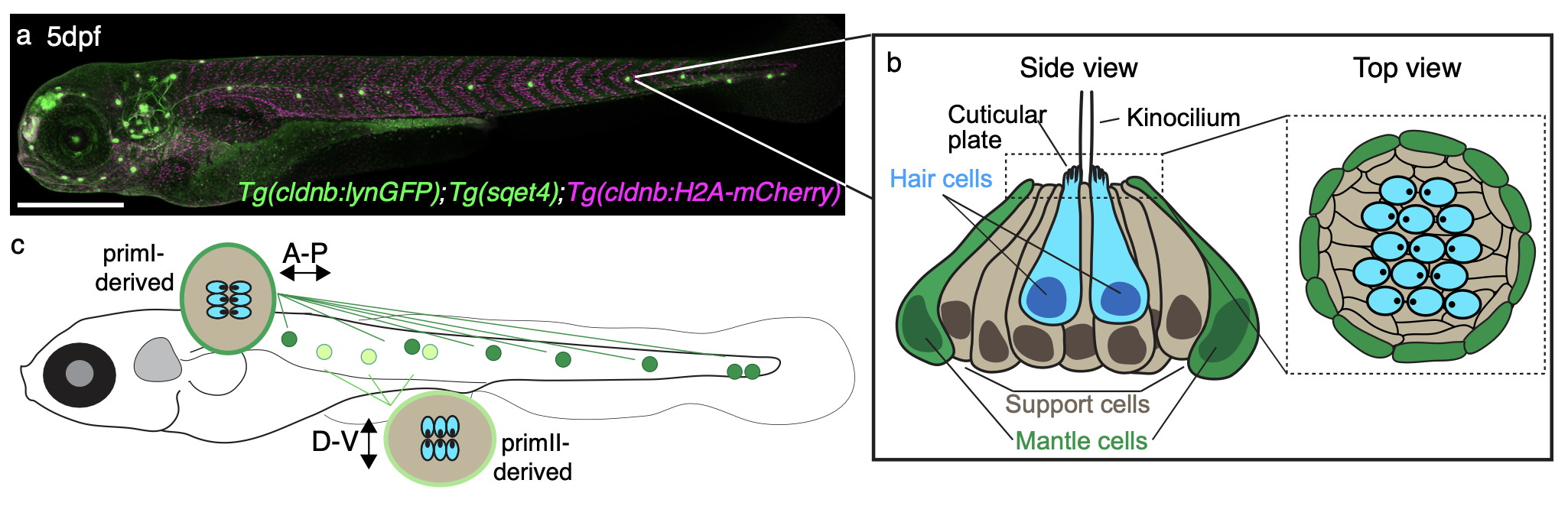
The video depicts the formation of the so-called lateral line in a transgenic zebrafish that I took when I was a student in Tatjana Piotrowski’s lab at the Stowers Institute for Medical Research (Kansas City, MO, USA). Cells of the lateral line use collective cell migration —sort of a cellular ‘Conga’ formation— to move from the head of the animal into the trunk and deposit volcano-shaped structures called neuromasts.
Where can the lateral line be found?
Fishes (bony, such as the zebrafish; and cartilaginous, such as sharks and rays) and amphibians (such as frogs and salamanders).
How was this video taken?
This is a video of a transgenic zebrafish expressing a fluorescent protein in the lateral line Tg(clndb:lynGFP). The video was taken live using a Zeiss 780 confocal microscope.
What does the lateral line do?
The lateral line is a mechanosensory organ that aquatic animals use to orient themselves in the water using neuromasts that cover the entire body of the animal (like in the picture below). These neuromasts are the sensory unit of the lateral line due to the presence of specialized sensory cells called hair cells (that do indeed have little ‘hairs’, but we call them kinocilia and stereocilia) that respond to the water flow. This movement is translated into synaptic information and is sent to the brain, where it is used to convey positional information. The lateral line also allows fishes to display the so-called ‘schooling behavior’, a kind of collective animal behavior that is commonly seen in documentaries showing fishes moving together
Why should people care about this?
Due to the transparency, rapid development and gene conservation of zebrafish, the lateral line is an outstanding model to study two processes: collective cell migration, and hair cell regeneration.
The migrating primordia depicted in the video deploy the same molecular and cellular tools healthy and cancer cells use to migrate. Thus, studying migration of the lateral line primordia can help us understanding collective cell migration in health and disease.
One interesting property about hair cells is that they are also present in humans, but they are in the inner ear and they are used for hearing. When we age, or under some non-physiological conditions, we lose hair cells forever. Fishes, on the other hand, can regenerate their hair cells upon loss; therefore, studying how fish regenerate their hair cells may give us clues that can be used to restore hearing in humans that have lost hair cells.
How would you explain this to an 8-year-old?
During their development, fishes have little groups of cells that move all together all over the body. These cells then form garlic-bulb shaped structures that fishes use to swim, due to the presence of prickly cells called hair cells. People also have hair cells — but not all over their bodies, but inside your ears — that we use for hearing.
Where can people find more about it?
In the video, a lateral line primordium migrates to the tip of the tail of a zebrafish embryo
In this post, I invite you to join me on the journey through our recent article titled “Two Orthogonal Differentiation Gradients Locally Coordinate Fruit Morphogenesis.” This story started when I joined the lab of Daniel Kierzkowski at the Institut de Recherche en Biologie Végétale (IRBV) at the Université de Montréal as a visiting PhD student in January 2020. My main goal was to learn how to perform live imaging in plants and analyze the output data. Daniel invited me to contribute to his project related to fruit morphogenesis, and I believe it fit perfectly my interest in fruit development. The central question of this project was straightforward: what are the growth patterns underlying gynoecium development from its initiation to the final shape? I started working on that project just one week prior to the COVID outbreak in Canada. I had my first confocal experiment running the day before the University of Montreal recommended all foreign internship students to go back home due to upcoming pandemics. I decided to stay and found myself “stuck” in Montreal. This situation led to my most productive period when I spent several hours, days and weeks in the microscopy room. It was just me and the confocal microscope in the entire building (a quiet solitary experience). Walking through the deserted corridors evoked a mix of sadness and loneliness, but there was also a rewarding feeling of the “perfect samples” I just imaged with the confocal microscope. We were all astonished to see that my samples could grow for two consecutive weeks, from small primordium to fully developed gynoecium (future fruit)!
Simultaneously, I juggled the demands of writing my PhD thesis, preparing one of my papers derived from my PhD, and gearing up for my thesis defense, which, due to the outbreak, had to be conducted online. It was a challenging time, but one that ultimately shaped me and my research journey in unexpected ways.
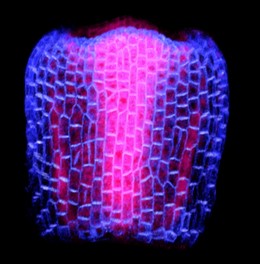
After successfully obtaining high-quality images of the gynoecium, we started the image analysis using MorphoGraphX. This part would not have been possible without the help of my colleague Elvis Branchini, whose dedication helped us segment and quantify growth in thousands of cells (from around 50 to 11000 per sample). This initial analysis represents, to our knowledge, the first comprehensive growth analysis spanning organ initiation to full expansion in plants. I take great pride in this significant achievement.
We started to dissect in detail different analyses on our gynoecium data including cell expansion, size, anisotropy, proliferation, and differentiation from each single cell. By deconstructing growth directions, both medial-lateral and longitudinal, we observed a medial-lateral growth gradient early in development. This observation contrasts with other plant organs, such as leaves, sepals, and stamen, which typically exhibit a basipetal (top-to-bottom) gradient of growth. This result was surprising for us: an organ that comes from an ancestral leaf has a different behavior. Then we started to look for explanations and formulated different hypotheses. Among these, the role of auxin — a favorite hormone in the plant scientific community — was promising within the context of our study.
Perspective from this Study:
From this study, we found that two distinct, time-shifted, and competing differentiation gradients govern gynoecium morphogenesis: an early mediolateral growth gradient and a late longitudinal growth gradient. A compelling next step would be to explore how these gradients interact. It would be fascinating to examine whether the early differentiation of the valve restricts the typical basipetal gradients from spreading through the organ, similar to what is observed in leaves, sepals, and petals. Additionally, investigating how the timing of these gradient establishments affects the final fruit shape in Brassicaceae, which exhibits a wide variety of fruit shapes, could provide valuable insights.
One of our favorite pictures, a gynoecium at 2 (2DAI) showing PIN-FORMED1 (PIN1), expression. PIN1 is a protein carrier and a key mediator in the transport of the plant hormone auxin. In the image, blue indicates PIN1 expression in the epidermal layer, while pink shows PIN1 expression in the internal layers.
My contribution:
The methods and approaches I developed for this paper are now being used in Daniel’s lab. These techniques will facilitate a more detailed investigation into fruit development across different species, improving our understanding of how fruits develop in the Brassicaceae family. This study offers a thorough and detailed atlas of growth patterns during gynoecium development. While many fascinating questions about fruit shape remain, this research paves the way for a deeper exploration of fruit development, particularly focusing on the shape and mechanical interactions within its different tissues.
My eureka moment:
Each step of the project felt like a eureka moment to me, but one has stayed with me: when we observed a full series growing continuously for two consecutive weeks. It was a delightful surprise. Additionally, each session of live imaging proved to be both gratifying and occasionally frustrating, yet undeniably worthwhile. My colleague and friend Binghan can attest to this, having shared the excitement, and participated in insightful discussions about our findings.
Bumps along the way:
Like many academic research projects, our journey with the gynoecium project was marked by challenges. We faced setbacks, moments of being stuck, and occasional frustration. I lost count of the numerous samples that did not survive or perished along the way. I recall one particular incident during my chemical treatment experiment. The plants were ready, I meticulously dissected numerous samples to maximize our chances of success. It was my fourth day of imaging, and the samples were growing really happily. Then, a disaster struck on a Sunday evening. I went to switch on the confocal microscope, and guess what? I could not initiate the system, it crashed, I wanted to cry at that moment (but I did not). Then, I lost my samples. I had to repeat the experiment again.
Similar situations occurred a couple more times, leading my colleagues to jokingly label me as having bad luck. However, I learned to cultivate resilience and approach failed experiments with a sense of humor —’Here we go, again!’— became my mantra in the realm of science.
Along the way, I learned invaluable lessons, the journey of trial and error ultimately led to new discoveries, making the effort worthwhile. When I look back and see everything we have made, I think it was all worth the effort! Despite the technical obstacles, I persevered, allowing me to expand my skills in problem-solving, critical thinking, and patience.
Our experience with the review process for our article submission was surprisingly smooth, especially compared to the tales I have heard from my colleagues. I have no complaints in that regard; both the editors and the reviewers were prompt in their responses. The comments of all the reviewers helped us improve our story. Finally, our story has found a home where it can be read.
My next step:
I have found myself profoundly inspired by this project. This experience has solidified my conviction that within science, limitless opportunities await those who approach their work with love, passion, and genuine curiosity. As scientists, we are not just observers; we are creators, empowered to innovate and explore the unknown. Following this project, I am eager to continue working on plant development and plant hormones. My focus will be on synthetic biology, and I aim to learn and utilize cutting-edge techniques such as single-cell sequencing, proteomics and CRISPR-Cas. I want to combine my knowledge of plant development and synthetic biology with one of my passions: microscopy. I’m excited to see what discoveries await on my next journey!
In the end:
The journey through this paper has significantly enhanced and refined several skills crucial for my scientific career. This accomplishment is deeply indebted to the invaluable assistance of my colleagues, who dedicated countless hours to segmenting hundreds of cells. Furthermore, the support of my friends and peers has been invaluable. Finally, I want to express my gratitude to my former boss, Daniel, for his ongoing support, insightful feedback, and discussions throughout my stay in his lab.
“A picture of the room where I spent hundreds of hours imaging, and my favorite tool, the microscope, which was used to build this story.”
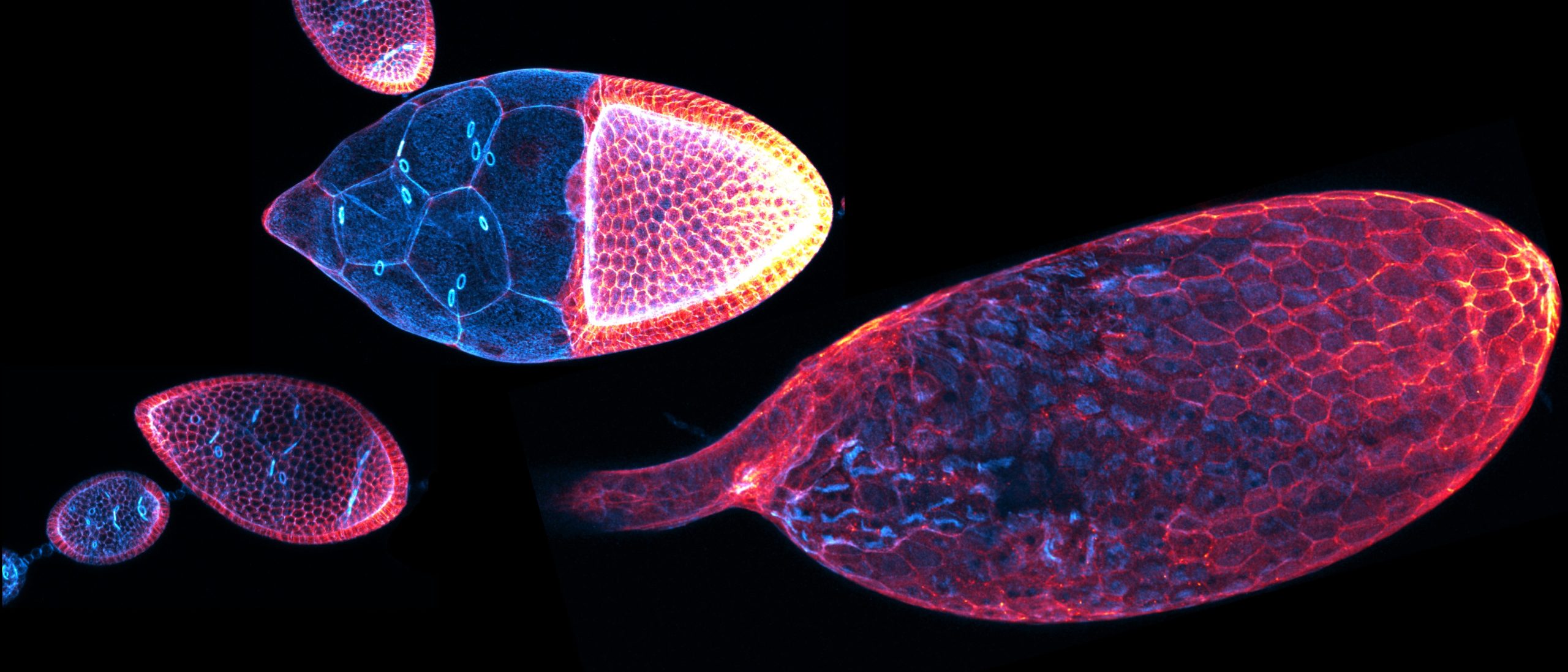
Basement membranes (BMs) are thin, specialized extracellular matrices that surround most tissues and organs (Jayadev and Sherwood, 2017). These meshworks serve as scaffolds for cell adhesion, influencing cell signaling, cell migration, proliferation, and differentiation (Sherwood, 2021; Yurchenco, 2011). Moreover, dysregulation of BM remodeling lead to disturbed tissue and organ development or disease (Sekiguchi, R and Yamada, K. M., 2018). Recent publications indicate that establishment of BM heterogeneity might be important for tissue and organ sculpting (Agarwal et al., 2022; Harmansa et al., 2023; Harunaga et al., 2014; Kyprianou et al., 2020; Serna-Morales et al., 2023; Uwe Töpfer et al., 2022). However, how this heterogeneity is induced and how this leads to organ sculpting is largely unknown.
In our recent publication (Töpfer et al., 2024), we identified two AdamTS matrix proteases required for the proper elongated shape of the egg chamber. Knockdown of stall or AdamTS-A results in rounder eggs from early elongation phase on. While the phenotypes look very similar, the molecular mechanisms by which they act are different. Using CRISPR/Cas, we tagged both proteins with sfGFP and found a dynamic expression, resulting in higher protein enrichment in the terminal regions (at the most anterior and posterior regions). We were able to detect Stall in early stalk cell precursors and stalk cells as well as a strong expression later in the polar cells with a gradual expression at the terminal regions of stage 8 egg chambers. AdamTS-A was uniformly expressed in all somatic precursors of follicle cells, but becomes more strongly enriched at the terminal regions in stage 8 egg chambers, too. Next, we used fly lines with GFP-tagged ECM components to study the proteases’ role in ECM remodeling. We found that Stall is required to establish basement membrane heterogeneity by locally limiting Collagen IV protein density. In contrast to a lower fluorescence signal in control egg chambers in the posterior region, stall knockdown led to a nearly uniform protein level of Collagen IV along the anterior -posterior axis. Using high-resolution microscopy, we studied the pattern of fiber-like structures embedded in the BM. We found that the knockdown of AdamTS-A results in a disturbed BM micropattern. BM fiber-like structures were shorter and smaller in AdamTS-A knockdown egg chambers. Maturation (length and proper orientation) of BM fiber-like structures has been associated with egg chamber rotation. Accordingly, we also found that AdamTS-A is required for proper egg chamber rotation, hence knockdown of AdamTS-A results in a premature stop. We performed Atomic force microscopy to measure the stiffness of the BM. In both knockdown conditions, BM stiffness was globally increased, what goes along with increased apical pSRC level. Finally, we found slower E-Cad recovery in a FRAP experiment and a disturbed cell aspect ratio in the central regions. This data indicates that basement membrane remodeling by AdamTS-A and Stall influences gradual BM remodeling which induces BM stiffness and cell shape globally, which is required for organ shape.
References
Agarwal, P., Shemesh, T. and Zaidel-Bar, R. (2022). Directed cell invasion and asymmetric adhesion drive tissue elongation and turning in C. elegans gonad morphogenesis. Developmental Cell57, 2111-2126.e6. Harmansa, S., Erlich, A., Eloy, C., Zurlo, G. and Lecuit, T. (2023). Growth anisotropy of the extracellular matrix shapes a developing organ. Nat Commun14, 1220. Harunaga, J. S., Doyle, A. D. and Yamada, K. M. (2014). Local and global dynamics of the basement membrane during branching morphogenesis require protease activity and actomyosin contractility. Developmental Biology394, 197–205. Jayadev, R. and Sherwood, D. R. (2017). Basement membranes. Current Biology27, R207–R211. Kyprianou, C., Christodoulou, N., Hamilton, R. S., Nahaboo, W., Boomgaard, D. S., Amadei, G., Migeotte, I. and Zernicka-Goetz, M. (2020). Basement membrane remodelling regulates mouse embryogenesis. Nature582, 253–258. Sekiguchi, R and Yamada, K. M. (2018). Basement Membranes in Development and Disease. Current Topics in Developmental Biology130, 143–191. Serna-Morales, E., Sánchez-Sánchez, B. J., Marcotti, S., Nichols, A., Bhargava, A., Dragu, A., Hirvonen, L. M., Díaz-de-la-Loza, M.-C., Mink, M., Cox, S., et al. (2023). Extracellular matrix assembly stress initiates Drosophila central nervous system morphogenesis. Developmental Cell58, 825-835.e6. Sherwood, D. R. (2021). Basement membrane remodeling guides cell migration and cell morphogenesis during development. Current Opinion in Cell Biology72, 19–27. Töpfer, U., Guerra Santillán, K. Y., Fischer‐Friedrich, E. and Dahmann, C. (2022). Distinct contributions of ECM proteins to basement membrane mechanical properties in Drosophila. Development149 (10): dev200456. Töpfer, U., Ryu, J., Guerra Santillán, K. Y., Schulze, J., Fischer-Friedrich, E., Tanentzapf, G. and Dahmann, C. (2024). AdamTS proteases control basement membrane heterogeneity and organ shape in Drosophila. Cell Reports43, 114399. Yurchenco, P. D. (2011). Basement Membranes: Cell Scaffoldings and Signaling Platforms. Cold Spring Harbor Perspectives in Biology3, a004911–a004911.


 (3 votes)
(3 votes)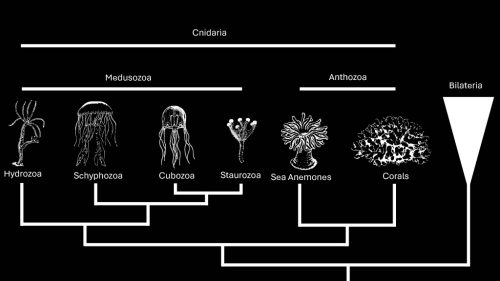
 (No Ratings Yet)
(No Ratings Yet)