A postdoctoral research associate position is available immediately to investigate the role of reactive oxygen species (ROS) as physiological signaling molecules in axonal growth and guidance in the lab of Dr. Daniel M. Suter at Purdue University, West Lafayette, IN https://suterlab.bio.purdue.edu/. Specifically, the work will investigate the molecular and cellular mechanisms of how ROS regulate axonal growth and guidance in vitro and in vivo. The successful applicant has experience in cell biology and molecular biology. Additional expertise in developmental neuroscience, microscopy, and working with zebrafish is desirable but not required. This position is supported through funding from the NIH.
Highly motivated candidates with a PhD in cell biology, molecular biology, or neuroscience who are interested to participate in this exciting project at the interface of cellular and developmental neurobiology are invited to send their CV, a brief statement of research interests and contact information of three referees to Dr. Suter.
Dr. Daniel M. Suter
Department of Biological Sciences
Purdue University
915 West State Street
West Lafayette, IN 47907
USA dsuter@purdue.edu
765-496-1562
The BiOf lab http://biof-lab.org/ has developed a microfluidic technique, the Cellular Capsules Technology, that allows them to produce multicellular spheroids and organoids in a high throughput and controlled format. The applications in tissue engineering, oncology and regenerative medicine and toxicology testing are numerous. The recruited postdocs will be involved in the interdisciplinary projects pursued by the team in engineering multiscale vascularized tissues, reconstructing a functional liver lobule, deciphering the impact of mechanical cues on hepatocarcinoma and glioblastoma progression in vitro models, developing innovative microscopy techniques for thick tissue imaging.
We seek for talented scientists with exceptional motivation and outstanding expertise (i.e. PhD) in biophotonics /image analysis, cancer or stem cell biology, microfluidics, biophysics or tissue engineering.
The proposed funding is for 12 months and can be renewed up to 36 months. The salary will be adapted to the experience of the applicant. Starting date will be between March and June 2021.
A postdoctoral research position is available in the group of Dr. Kristen Panfilio at the University of Warwick, UK, to join our BBSRC-funded project on how polyploid nuclear structure influences cellular function in dynamic epithelial tissues. This is a full-time, fixed-term position for up to 36 months, integrating developmental biology, bioinformatics, cell cycle regulation, genome organization, and 4D live cell imaging.
Polyploidy is hypothesized to aid epithelial barrier formation and its repair after wounding and to rapidly supply gene products by transcription from multiple gene copies. Yet polyploid genomic structure may be unstable and require active inhibition of apoptosis through regulatory processes that are not yet well understood. Investigating the costs and benefits of polyploidy is essential to understand tissue-specific development, homeostasis, and ageing.
The two extraembryonic tissues of insects offer an excellent – and inherently comparative – research model, spanning key developmental stages for morphogenesis and cellular physiology. Our research species is the flour beetle Tribolium castaneum, which offers advanced genetic resources and where we integrate fluorescent live cell imaging and next-generation sequencing approaches (see the lab’s recent publications in eLife 5:e13834, Development 143:3002, Commun. Biol. 3:552). The project uses methods to assess nuclear size and tissue integrity, quantify gene expression, and genetically challenge barrier organization and cell number. Altogether, we will test long-standing hypotheses on polyploidy function and its end-stage implications in animal tissues. A complete description of the project is available at: https://gtr.ukri.org/projects?ref=BB%2FV002392%2F1.
You will have a Ph.D. or equivalent and good knowledge and experience in genetics, cell and developmental biology, standard molecular biology techniques,and working with NGS data, as evidenced by your Ph.D. thesis and/or authored papers in peer-reviewed journals. Familiarity with the fields of insect developmental genetics and comparative genomics would also be highly advantageous. Practical experience in any of the following is desired: advanced microscopy (including light sheet fluorescence microscopy),RNAi, FACS, RT-qPCR, and computational work with either sequencing or imaging data. We will provide full training in new techniques, supported by the possibility for international travel and collaboration.
Enquiries and expressions of interest directly to Kristen (K.Panfilio@warwick.ac.uk) are welcome, with applications made on-line (post number 103508). Full applications will include contact details for at least two referees, a CV, and covering letter stating why you are interested in the topic and what you would bring to the project. The application closing date is 7th February 2021.
The patterning of stomata – the pores in the plant epidermis that facilitate gas exchange and water control – is regulated by a family of small secreted peptides. A new paper in Development analyses the effective ranges of two such peptides, borrowing a statistical technique used by astrophysicists to investigate the distribution and patterning of galaxies. We caught up with authors Emily Lo, who worked on the project when an undergraduate at the University of Washington (UW), and her supervisor Keiko Torii, who recently moved her lab from UW to The University of Texas at Austin (where she is Professor and Johnson & Johnson Centennial Chair in Plant Cell Biology), to hear more about the story.
Emily (L) and Keiko (R)
Keiko, can you give us your scientific biography and the questions your lab is trying to answer?
KT: As a plant developmental biologist, I was always super fascinated by how dazzling arrays of functional, beautiful patterns emerge during development and how the external environment, where plants thrive, shapes the manifestation of such functional patterns. My main focus is to tackle such observations through understanding the molecular and genetic basis of cell-cell communication, ‘how plant cells talk to each other to generate functional patterns’.
I received my PhD and did my first short postdoc in Japan, where I identified the causal gene for the Arabidopsis mutant, erecta, which exhibits short stature and altered organ shape. It turned out that the ERECTA gene encodes a putative receptor kinase (collectively known as ‘Receptor-like Kinase’ or ‘RLK’), and it was the first report that this type of putative peptide receptor regulates plant growth and development. Later, after I obtained a tenure-track Assistant Professor position at the UW in Seattle, our research elucidated that ERECTA-family RLKs perceive a family of peptides to enforce proper stomatal patterning, which is the basis of this work.
In the Fall of 2019, I accepted the Johnson & Johnson Centennial Chair in Plant Cell Biology at the Department of Molecular Biosciences, The University of Texas at Austin. Currently I am also a Howard Hughes Medical Investigator.
And Emily – how did you come to work in Keiko’s lab on this project?
EL: I was lucky enough to be hired as an undergrad in Keiko’s lab during my freshman year at UW, and I stayed on until I graduated. It’s a rare opportunity to have such a long research experience as an undergrad, so I was able to devote a substantial amount of time to this project. Keiko introduced me to this fascinating project using mosaic fluorescent sectors to track peptide expression, which had been initiated by Dr. Takeshi Kuroha, a previous postdoctoral fellow, and Janelle Sagawa, a previous undergrad researcher (both acknowledged in the manuscript). So right away I could start growing seedlings, generating these mosaic sectors, and imaging on the confocal microscope. It was great fun! I graduated before we were able to complete the project, so Scott [Zeng] developed the majority of the SPACE pipeline after I left the lab.
How has your research been affected by the COVID-19 pandemic?
KT: Unfortunately, my lab and research program got hit really hard by the COVID-19 situation, because we had just relocated from Seattle WA to Austin TX right before the pandemic hit. Our brand-new lab at UT Austin was finally operating in the beginning of 2020 when we had to suddenly shut it down. Like myself, almost all the new lab members were new to Texas (or the Southern part of USA), and many were separated from family. So, staying at home in an unfamiliar city was stressful to everyone, and I truly thank my lab members for being positive and hanging together during this exceptionally difficult time. As for this manuscript, since we were at the phase of drafting a manuscript (lucky us!), Scott and I completed the manuscript during the full shut down phase, with thoughtful inputs from Emily and other co-authors. New online technology, such as Zoom, helped us work together, remotely.
EL: For this project, it was fortunate in that all experiments were completed and we were at the data analysis phase before COVID hit, so we were able to complete it through online communication (and because I’m located in Baltimore, I’d been communicating solely online anyway). For my current research at Hopkins, we were in an Essential-Only phase from March to June, in which no new experiments were allowed to begin; luckily, at that point I did have some computational analysis to catch up on. In mid-June we had our Phase-I reopening, so I’ve been able to resume many of my experiments, though of course progress is still limited by how much time we’re allotted in the lab.
Why have the signalling ranges of peptides like EPF1 and Stomagen been hard to assess, prior to your paper?
EL & KT: Whether secreted peptides or small chemical hormones, how far the signal moves is a fundamental question of pattern formation in development. But precisely quantifying the distance is not trivial. For direct observation, one could tag fluorescent proteins (or exogenously synthesize a peptide conjugated with a fluorophore). In a strict sense, however, such modifications change the size and property of peptides or chemical signals. Alternatively, one could develop sensors (such as a FRET sensor) that detect the existence of peptides or chemical signals.
We have previously shown that EPIDERMAL PATTERNING FACTOR (EPF) family members of secreted peptides fine-tune patterning of stomata on the plant epidermis by competitive binding to the same receptor. Because of the flat, two-dimensional nature of the developing leaf epidermis, we thought that our system would be a great model for understanding how far the secreted peptides influence tissue patterning, rather than directly observing its movement.
Can you give us the key results of the paper in a paragraph?
EL & KT: Using recombination-based mosaic sectors to overexpress signaling peptides EPF1 and Stomagen, which inhibit and promote stomatal development, respectively, we determined the effective ranges of these peptides in Arabidopsis cotyledons. We developed a quantitative pipeline to model stomatal distributions across the cotyledon in response to peptide overexpression, which we named SPACE (stomata patterning autocorrelation on epidermis), an homage to the astrophysics origin of the autocorrelation approach. We found that the inhibitor peptide EPF1 has a longer effective range than the activator peptide STOMAGEN, and that the patterning effects of peptide overexpression are limited to a local range rather than the global cotyledon.
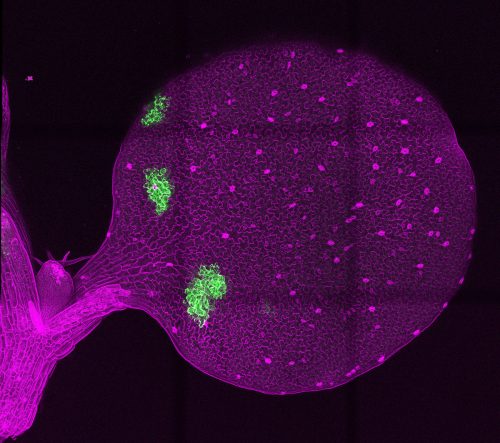
Tile scan of Arabidopsis thaliana cotyledon with mosaic sectors simultaneously overexpressing GFP, retained in the endoplasmic reticulum, and secreted peptide EPF1, an inhibitor of stomatal differentiation.
How did you come upon the idea of borrowing an astrophysical technique to look at stomatal patterning?
KT: At the initial stage of our research, we were able to produce chimeras via heat-shock Cre/lox recombination, but we could not think of how to actually ‘calculate’ the effective distance of the peptides. Initially, we tried to set bin range (such as 100 μm, 200 μm, etc.) from each sector border. However, because of the complex geometry of pavement cells as well as the unique size, shape and location of individual sectors, we could not figure out how to normalize stomatal distribution surrounding each sector.
One morning in the kitchen, I was talking about this problem to my spouse, who is a theoretical physicist studying String Theory. When I was drawing a cartoon of a simple leaf (essentially an oval) with lots of stomata (essentially dots inside the oval), he told me, ‘this sounds familiar to me. I think my colleague, Miguel, is addressing the exact same problem – except that in his case, it’s the distribution of galaxies in the Universe’. I immediately e-mailed Prof. Miguel Morales of UW Physics. I thought that he might think I was crazy, but to my pleasant surprise, Miguel and his postdoc Dr Bryna Hazelton were very excited to hear about our research and the potential of the spatial autocorrelation statistics that they utilize for astrophysics in solving questions in plant development. Bryna mentored Emily in programming for the spatial autocorrelation analysis.
My Physics colleagues generously gave me the opportunity to give a 15 min talk to incoming Physics Graduate Students. I discussed the principles of spatial patterning in biological systems and introduced Alan Turing’s reaction-diffusion model; luckily, Scott remembered my brief talk and was curious enough to join my lab to tackle this problem after Emily’s graduation.
EL: For me it started when Keiko suggested we meet with Miguel and Bryna. When we explained that we were trying to generate a metric of epidermal spatial patterning, they almost immediately suggested looking into autocorrelation, an astrophysics method for analyzing galaxy distributions/patterning. Bryna works at the UW eScience Institute, and has a goal to empower the next generation of researchers and students to answer fundamental questions in complex or noisy data. Working with her to develop code to analyze our epidermal patterning questions was a really wonderful and fruitful experience, and that was the starting point for our collaboration and the development of the SPACE pipeline.
Why do you think the stomatal inhibitor EPF1 is able to travel farther than the activator Stomagen?
EL & KT: To clarify your question, the goal of our study was to determine how far these peptides act or have an effect, not necessarily the physical distance they travel. EPF1 has a longer range in terms of developmental outcome than Stomagen, but not necessarily diffusion distance. The question of their differing ranges is complex because stomatal lineage cells (which are induced by Stomagen) will themselves secrete EPF1, acting as a negative-feedback loop; in other words, the peptides are not acting in isolation. Our quantitative determination of peptide effective range is fascinating because in the Turing theory of pattern formation, a short-range activator often interacts with a longer-range inhibitor to achieve a self-regulating, periodic pattern. The relationship between Stomagen and EPF1 might be one such example of this effect to achieve regular, ordered stomatal spacing across the epidermis.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
EL: After we’d established that the mosaic sector generation system indeed worked, we wanted to observe the potential global effects of peptide overexpression (from within the restricted mosaic sectors) across the entire cotyledon, so I switched to a confocal microscope that had tile-scanning functionality. The stitched image I got of the entire cotyledon surface was very beautiful; getting that first image was a hugely satisfying and validating moment for me.
Getting that first image was a hugely satisfying and validating moment for me
And what about the flipside: any moments of frustration or despair?
EL: The initial months were the most difficult experimentally for me. There were a lot of experimental skills that required finesse, for example sowing tiny Arabidopsis seeds individually on plates or mounting the cotyledons on glass slides completely flat without folding or tearing, etc. The way I overcame these challenges was practice and repetition: those are just skills that are gained slowly over time.
I understand you’ve now left Keiko’s lab – what are you doing now?
EL: I’m currently pursuing my PhD in Biomedical Engineering at Johns Hopkins, in the labs of Prof. Patrick Cahan and Prof. Andrew Feinberg. I study how changes in cell identity relate to cancer initiation in the context of pancreatic ductal adenocarcinoma: one of the most lethal malignancies in the US.
Even though I work on human disease research now, plant research is still very important to me, especially given the adaptations humans will have to make to agricultural practices in response to global climate change. Particularly, I think optimizing crop water consumption will be a key strategy in the next several decades to reach necessary agricultural yields in fluctuating environmental conditions.
Where will this story take the Torii lab?
KT: Stomatal patterning and distribution is critical for plant productivity and water use efficiency, and many different genetic and environmental factors (such as temperature, light, CO2 and drought) influence stomatal number, density and distribution. There is a long history in Plant Physiology of studying these traits. Yet, essentially, there are two ways to quantify these traits: stomata density (number of stomata per given area) and stomatal index (number of stomata per total epidermal cells). Different genetic backgrounds and environmental conditions may influence number, density or distribution in unique ways, but often such information is lost by simply presenting bar graphs of stomatal density and index. So, the natural next direction is to apply SPACE pipelines to describe different patterns of stomata, in different ages of Arabidopsis leaves or under different genetic/environmental conditions. Further harnessing the SPACE pipeline to quantitatively characterize stomatal patterning of agronomically important plant species, such as tomato and cereals, may reveal some important characteristics. Finally, we are looking for developmental biologists studying pattern formation of any systems who are brave enough to try out our SPACE pipeline to quantitatively characterize their systems.
Finally, let’s move outside the lab – what do you like to do in your spare time in Baltimore and Austin?
KT: Well…since I just relocated alone to a small one-bedroom apartment in Austin, and then the COVID-19 stay-at-home order was put in place, I really did not have much time to explore my new city. My spouse and children were supposed to move to Austin from Seattle this summer, but this got postponed due to COVID-19. I really hope that, when we (the USA) manage to get the pandemic under control, I can explore the Texas Hill Country and enjoy its natural beauty, and see bluebonnet wildflowers blooming for the first time in my life!
EL: I’m an avid baker, cook and gardener. During COVID-19, I’ve boarded the sourdough starter bandwagon as an at-home quarantine activity. And as much as possible, I’ve tried to stay in touch with friends and family across the country and the world by video chatting.
In the developing spinal cord, progenitor cells sequentially give rise to motor neurons and precursors of one of the major glial cell types: oligodendrocytes. A new paper in Development unpicks the molecular control of the neuron-glia switch and the differentiation of oligodendrocyte precursors in the zebrafish embryo. To find out more about the work, we met first author and graduate student Kayt Scott and her supervisor Bruce Appel, who holds the Diane G. Wallach Chair of Pediatric Stem Cell Biology and is Professor and Head of the Section of Developmental Biology at the Department of Pediatrics, University of Colorado School of Medicine in Aurora.
Kayt (L) and Bruce (R).
Bruce, can you give us your scientific biography and the questions your lab is trying to answer?
BA: I earned my PhD at the University of Utah under the mentorship of Shigeru Sakonju. My thesis work focused on transcriptional control mechanisms in the context of homeotic genes and developmental patterning, and it was during that time that I became interested in problems of cell fate specification. I then moved to the University of Oregon for postdoctoral training with Judith Eisen. Using zebrafish embryos, Judith had previously performed a brilliant set of experiments that led her to conclude that communication between newly born motor neurons determined their subtype identities. As I worked to try to figure out how those subtype identities were specified, other work in the field began to suggest that the same progenitor population that produces motor neurons also gives rise to oligodendrocytes: the myelinating glial cell type of the CNS. I initially thought oligodendrocytes and myelin were pretty boring but as we developed new tools and learned how to do time-lapse imaging, we discovered that these cells and their ability to form a myelin sheath on an axon is incredibly fascinating. This has spun off into many projects investigating the molecular and cellular mechanisms that regulate developmental myelination and how myelin changes in response to brain activity. However, I still consider the age-old question of cell fate specification to be the foundation of all that we do.
And Kayt – how did you come to work in Bruce’s lab and what drives your research today?
KS: Developmental biology has always been a passion of mine, especially in the context of early nervous system formation. So, before I had even applied to graduate school, I was aware of and intrigued by the work conducted in Dr Bruce Appel’s lab. In fact, I discussed my interest in his research in my application letter. Once I entered graduate school, I did a rotation in Dr Appel’s lab and began the study of Prdm8 function in the zebrafish spinal cord. By the end of my rotation, I was sure I wanted to be part of the lab and further my rotation project. Through my experiences in the lab, I have really become interested in understanding how early tissue structure is organized and how this organization is altered through time and these questions are what drives my research today.
How has your research been affected by the COVID-19 pandemic?
KS: At the time of the pandemic shut down I was working from home already, feverishly compiling this manuscript and working on a fellowship application, so the first couple of months weren’t too disruptive. However, after accomplishing these endeavours, I have been unable to get into the lab and further explore the mechanisms of how Prdm8 regulates Shh signalling response. Fortunately, this has provided me with a unique opportunity to delve into the single cell RNA-seq dataset we had generated and develop my bioinformatic skills. This has opened many new avenues of research in the Appel lab.
I am hopeful that the pandemic is making us more collaborative and communicative
BA: I am hopeful that the pandemic is making us more collaborative and communicative. Although we all miss interacting with our friends, colleagues and trainees, the pandemic is forcing us to find new ways to share information. I have been really impressed by how well the virtual scientific conferences are working. The conferences are now available to so many people who couldn’t afford them or had other barriers to travel. Additionally, distance no longer limits with whom we can share seminars or journal clubs or lab meetings. Consequently, I’m optimistic that our research will actually benefit because we will have better information and shared expertise.
In your efforts to understand the regulation of pMN development, what made Prdm8 a good candidate?
KS: Originally our interest in Prdm8 was motivated by understanding how oligodendrocyte precursor cells (OPCs) are maintained into adulthood. Prdm8 was identified in our bulk RNA-seq data as being highly expressed by OPCs and not expressed in oligodendrocytes, and because Prdm8 was a known transcriptional repressor that can complex with bHLH transcription factors, we predicted that Prdm8 was forming a complex with Olig2 to repress oligodendrocyte differentiation. As we began our investigation of Prdm8 function, we came across some interesting and unexpected phenotypes that transitioned our interests in Prdm8 function within pMN progenitors.
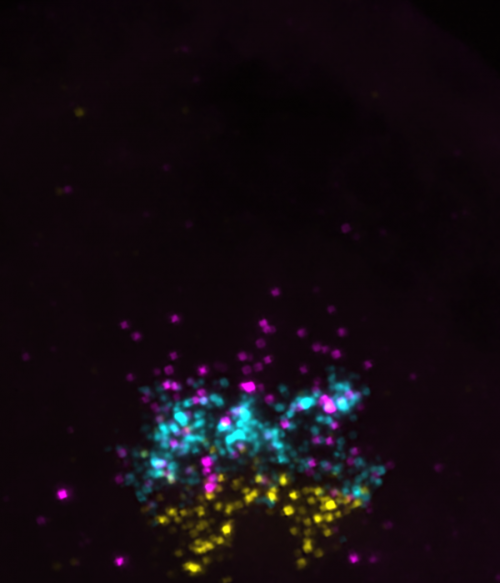
Transverse trunk spinal cord section showing olig2 (cyan), prdm8 (magenta) and nkx2.2a (yellow) mRNA at 24 hpf.
BA: Whereas much of the myelin field focuses on how oligodendrocytes make myelin, we have been curious about why so many OPCs persist without differentiating. Prdm8 jumped out of our RNA-seq data because OPCs expressed it at very high levels compared with myelinating oligodendrocytes. Because Prdm8 can function as a transcriptional repressor, the initial hypothesis was obvious. As Kayt notes, however, her work revealed that Prdm8 has an earlier function in regulating progenitor fate specification.
Can you give us the key results of the paper in a paragraph?
KS: We set out to understand the potential function Prdm8 plays in pMN cell fate. Detailed expression analysis revealed that pMN progenitors express prdm8 through development and that differentiating oligodendrocytes downregulate prdm8 expression, whereas OPCs maintain expression. We then learned that prdm8 mutant embryos have an excess of oligodendrocytes and a deficit in motor neurons as a result of a premature motor neuron-oligodendrocyte fate switch. This premature switch in cell fate was accompanied by a premature increase in nkx2.2a expression and ventral neural tube Shh activity with Prdm8 loss of function. By suppressing Shh signalling in prdm8 mutants, we were able to restore the motor neuron population but were not able to reduce the excess oligodendrocyte phenotype observed in mutants. In all, these data support the hypothesis that Prdm8 prevents the neuron-glia switch in pMN progenitors by suppressing Shh signalling activity, and, independently of Shh activity, Prdm8 regulates the balance of OPCs and oligodendrocytes.
Do you have any clues as to how, at the molecular level, Prdm8 inhibits Shh signalling?
KS & BA: One possibility we have entertained is that Prdm8 might form a complex with Olig2 to suppress transcription of genes that promote cellular response to Shh. Figuring this problem out will be the next big push for the Prdm8 project.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
KS: I would have to say the most striking result I came across in this project was that Shh repression was only able to rescue motor neuron populations, but resulted in a total oligodendrocyte lineage deficit while maintaining excess oligodendrocytes in mutant embryos. This was the point where it really sunk in that Prdm8 function was more complex than I imagined, and appeared to have temporal and spatially distinct functions even within a single lineage.
And what about the flipside: any moments of frustration or despair?
KS: The most frustrating part of this project was that I had started it as a rotation student and hadn’t quite honed my organizational skills. This resulted in having to redo several of the first experiments, because I had not kept good enough documentation of my methods or results. A lesson that will never leave me!
What’s next for you after this paper?
KS: Moving forward, I have been working on a scRNA-seq paper, characterizing pMN cell populations through time and have plans to graduate in the summer of 2021. Following graduation, I hope to obtain an IRACDA fellowship to receive comprehensive training in undergraduate education and develop my own research projects in the field of developmental biology.
Where will this story take the Appel lab?
BA: There is an old joke that, in the absence of good ideas, do a genetic screen. A modern-day version of that is to do single cell RNA-seq. We have been stumped about the earliest molecular events that specify neural progenitors for oligodendrocyte fate, so we carried out a series of single cell RNA sequencing experiments. The data are incredibly exciting and appear to match our previous fate-mapping studies suggesting that motor neurons and oligodendrocytes arise from distinct progenitors. We now have many new ideas to pursue in our attempt to gain a comprehensive understanding of the gene regulatory network that specifies oligodendrocyte fate.
Finally, let’s move outside the lab – what do you like to do in your spare time in Colorado?
KS: Outside of the lab, I never waste an opportunity to enjoy the beautiful state we live in. You will find me camping, hiking and fishing almost every weekend during the warmer months; in the winter, I am hitting the slopes on my snowboard or gliding through the woods on some cross-country skis.
BA: Now that our daughters are pretty much on their own, my wife and I try to spend as much time as possible at our place in the mountains. It’s off-grid and fairly remote, and a great jumping-off spot for hiking, backpacking, skiing and snowshoeing. I am also very slowly and ineptly trying to restore a 1942 Ford tractor that my parents had on their farm.
Welcome back to #DevBiolWriteClub. Let’s review the rules:
Do the work.
Do the work.
Revise and edit. Again, and again, and again.
Read with intent.
You can’t do it alone.
Last time, I hammered away at Rule #4, Read with Intent. Today, I’ll return to Rule #4, and I’ll make some preliminary comments on Rule #5. Rule #5 is mostly about being brave enough to show your writing to your peers, and even your mentors. But it’s also about taking advice where you can get it.
Thankfully, you are not alone in your struggles with writing. I’m right there with you, and so are hordes of other aspirants. So many, in fact, that there’s a brisk market in books about writing. So, let’s look at this: If you take the time to carefully read a book (or two, or three) about writing, you are hitting rules #1, #2, #4, and #5. Wow.
So, because books are always the best Christmas presents, here are #DevBiolWriteClub’s favorite books about writing.
If you’re only going to read one book about writing, read:The Scientist’s Guide to Writing, by Stephen Heard. This is unquestionably the best book for science writing. Stephen is a biologist, and he’s a delightful presence on Twitter (@StephenBHeard). He also has an awesome blog, where he frequently writes about writing. His book has everything: A brief and fun history of science writing, some big picture psychology at the start, then excellent “brass tacks” advice on writing scientific papers. I recommend this one especially to anyone following #DevBiolWriteClub and looking for new ways to “do the work.” Each chapter ends by suggesting helpful exercises.
If you write, but don’t really like what you write, read: Writing Science in Plain English, by Anne Greene, also a biologist. This very slim book (the core of it is 85 short pages) will likely do more to improve the sentences and paragraphs you write over the short term than any other book I’ve read. It’s a straight-up style manual focused on how to turn your scientific writing into simple prose and thus to communicate more effectively. Not surprisingly, this book’s simple, actionable advice is crystal clear. It also provides a series of short exercises to drive home the key points.
If you have trouble getting that first draft written, read: How to Write a Lot, by Paul Silvia. This is another little book, but it’s all about the big picture. Silvia is an academic psychologist, and his book is aimed at fixing bad habits of mind and creating new ones. Since scientist’s intellectual lives are complex enough already, an important asset is that while Silvia’s book attacks very high level issues, it offers remarkably simple and practical advice on improving your practice as a writer, the core goal of #DevBiolWriteClub. One of the best aspects of this book is an explicit description of various types of writing groups that help to address subtly different hurdles that writers may face. This really helps with Rule #5. This one is also just a fun read.
Ok, those are the Big Three, and any scientist wanting to improve their writing should read all three of these books.
Of course, we can always improve more, so here are some more recommendations:
The Sense of Style, by Steven Pinker. I read somewhere -possibly in this book- that every writer should read a style manual once per year, whether they need to or not. If you’re a working scientist, that book probably should be Writing Science in Plain English, above. But if you want to dive in a little deeper, there are tons of great style books. The canonical style manual is Strunk and White’s Elements of Style, but with apologies to essentially all of my mentors, I find it too stuffy and rather boring. Pinker’s book, which he freely offers as an update to that classic, is far more fun to read. This quip from the prologue really spoke to me: “You can write with clarity and with flair, too.”
The War of Art, by Steven Pressfield. Ok, this one is really just a self-help book. But if you’ve read How to Write a Lot, above, and you still have trouble sitting down and doing the work, read this.
Writing Your Journal article in Twelve Weeks, by Wendy Laura Belcher. Ok, it’s hard to say I “like,” this 400+ page book, but it does offer something none of the others do: Step-by-step instructions on writing a paper in a reasonable, totally defined time frame. It presents clear goals each day and each week. If you are serious about a 12-week, self-taught crash course in writing a scientific paper, pick this one up.
Bird by Bird, Some Instructions on Writing and Life, by Anne Lamott. This gem of a book is really targeted for aspiring poets and novelists, but since poets and novelists are often good writers (duh), there’s a lot here for us scientists. Chapter Three is entitled “Shitty First Drafts.” Need I say more?
On Writing, A Memoir of the Craft, by Stephen King. Yes, that Stephen King. Writing advice from one of the most successful authors in history? Of course, you should read it. If you’re in a hurry, you can skip the autobiography and go right to the writing advice about 100 pages in. But then you’d miss learning about his hardscrabble upbringing and the depth of his struggles even as a successful writer. Writing is hard, reading this book will help you really know that.
Understanding brain circuit evolution at single-cell resolution using comparative connectomics and transcriptomics
A position for a postdoc is available in the Kebschull Lab at the Department of Biomedical Engineering at the Johns Hopkins School of Medicine in Baltimore, MD. We develop and apply cutting edge molecular and neuroanatomical tools to study how primordial circuits expanded in evolution to form the complex brains that exist today. We have a special focus on barcode sequencing-based high-throughput connectomics (BRICseq, MAPseq) and in situ sequencing, which we apply in the cerebellar nuclei and brain-wide in different vertebrates. Recent relevant papers include Kebschull et al. 2020 Science, Huang et al. 2020 Cell, Han et al. 2018 Nature, and Kebschull et al. 2016 Neuron.
Candidates must hold a PhD degree (or equivalent) in neuroscience, biomedical engineering, molecular biology, or a related field. The ideal candidate should also have some bioinformatics skills and be passionate about brain mapping and evolution. We particularly encourage applications from any underrepresented or minority group.
Our lab is located on the School of Medicine Campus of Johns Hopkins University, surrounded by world-class neuroscience and biomedical engineering labs. We are committed to establishing a first-class, stimulating, diverse, and equitable environment in our new lab to allow you to flourish, achieve your goals, and further your career.
Qualified applicants should send a letter describing their current and future research interests, their CV, and names and contact details for three references to kebschull@jhu.edu. More information is available on https://www.kebschull-lab.org/.
Journal covers celebrate the aesthetic achievements of researchers and showcase the beauty of science to the wider world. Here are all 24 of Development’s covers for the crazy year that was 2020 – click the images to open the gallery, and you’ll find the cover description. Be sure to check out Development’s review of 2020 over on Twitter.
Which one’s your favourite? Leave a comment and we’ll let the authors know!
Micrographs of C. elegans anchor cells (cdh-3>mCherry::moeABD, magenta) atop the uterine-vulval basement membrane (laminin::GFP, green), expressing endogenously-tagged transcription factors (nuclear, green), grouped by column (left to right: egl-43::GFP, fos-1::GFP, hlh-2::GFP and nhr-67::GFP). Each row represents a different RNAi perturbation [top to bottom: empty vector, egl-43(RNAi), fos-1(RNAi), hlh-2(RNAi) and nhr-67(RNAi)]. See Research article by Medwig-Kinney et al. (dev185850)
Coronal section of a postnatal day 0 mouse brain stained to reveal EGFP from Dlx5/6-CIE cells in green, calbindin in red, and Hoechst in blue. Intact JNK signalling is required for Dlx5/6-CIE-positive interneurons to travel in tangentially oriented migratory streams to their correct positions in the developing cortical wall. See Research article by Myers et al. (dev180646).
Mouse primary cortical neurons cultured for 8 days in microfluidic chambers, which allow the compartmentalisation and fluidic isolation of axons from the cell bodies. Tau-1 immunolabelling (magenta) shows axons crossing via microgrooves into the axon compartment. Immunolabelling of the dendritic marker Map2 (green) shows dendrites and cell bodies restricted to the somatodendritic compartment. See Research article by Lucci et al. (dev180232).
An adult of the basal echinoid (cidaroid) Prionocidaris baculosa. The endomesoderm gene regulatory network (GRN) in cidaroids appears to preserve the ancestral state of echinoids and provides a valuable model for understanding the diversification of the early developmental GRN. See Research article by Yamazaki et al. (dev182139). Picture credit: Makoto Urata (Kanazawa University).
Dorsal view of the abdominal epidermis of a Drosophila pupa stained for E-cadherin to visualize cell junctions (white). The small histoblasts (cyan and black) proliferate and replace the large larval epidermal cells. Larval epidermal cells of one segment are coloured according to their position at the time of their extrusion: at the border of the histoblast nest (yellow), at the dorsal midline (green) or in between (magenta). See Research article by Michel and Dahmann (dev179606).
Artistic rearrangement of leaf confocal micrographs displaying battery of fluorescently tagged cell fate determinants in response to injuries. Images created by Anju P. S., Dhanya Radhakrishnan and Abdul Kareem V. K. Artwork by Mabel Maria Mathew (Indian Institute of Science Education and Research, Thiruvananthapuram, India). See Research article by Radhakrishnan et al. (dev185710).
Metatarsal from an E16.5 Lifeact-EGFP embryo (Lifeact is false coloured in pink, centre), which has been dissected and put into culture. Blood vessels (visualised with CD31, purple) sprout from the metatarsal to form a complex vascular network, allowing for ex vivo analysis of vascular density and sprout dynamics. See Research article by Schimmel et al. (dev185405).
Dorsal view of a Drosophila larval posterior segment, showing the topology of internal organs. The alary muscles (red) connect to the heart (phalloidin, blue; cardioblast nuclei, green) and to pericardiac cells (green) via an extracellular matrix network (pericardin, cyan) and maintain the heart and trachea (cyan) in proper positions. See Research article by Bataillé et al. (dev185645).
Polarising region grafts made to the anterior margin of a wing bud of a host chick embryo produce mirror-image duplications of the feather bud pattern, as shown by the black pigmentation. See Research article by Busby et al. (dev188821).
Proliferating Muller glia-derived progenitor cells in the chick retina. A vertical section of the retina was labeled for Sox2 (red), neurofilament (green) and phospho-histone H3 (blue). Inhibition of NF-κB-signaling stimulates the formation of proliferating Muller glia-derived progenitors. See Research article by Palazzo et al. (dev183418).
Volume rendering of an E16.5 mouse foetus based on X-ray microCT imaging. A novel protocol for staining cartilage matrix with Ruthenium Red is introduced that allows isotropic 3D imaging of the entire developing skeleton at micron resolution. See Research article by Gabner et al. (dev187633).
Line art traced using a real zebrafish retina image for reference and filled with single-cell transcriptome-defined clusters of cells depicted in different colours, representing a unified developmental program shared by embryonic and post-embryonic retinogenesis. See Techniques and Resources article by Xu et al. (dev185660).
Caenorhabditis elegans embryos imaged using single-molecule FISH to detect maternally-loaded mRNAs (clu-1, chs-1, imb-2, erm-1, F40G12.11, lem-3, nos-2 and cpg-2) in combination with cellular markers (PH::GFP, GLH-1::GFP, and DAPI). mRNAs accumulate at cell membranes, nuclear peripheries and within P granules. See Research article by Parker et al. (dev186817).
Root growth trajectories of Arabidopsis mutants in genes encoding RNA-binding proteins with specific N6-methyladenosine (m6A)-binding capacity, showing that a set of m6A-binding proteins controls directionality of root growth, and is required for rapid cellular proliferation during organogenesis, including root formation. See Research article by Arribas-Hernández et al. (dev189134).
Drosophila melanogaster embryos expressing an endogenously tagged EGFR-sfGFP construct showing localization of the EGFR in the developing tissue. Stage 7 (top), 9 (middle) and 11 (bottom) embryos are shown in a lateral view and are co-stained with EGFR-sfGFP (anti-GFP; green) and diphosphorylated ERK (dpERK; blue) antibodies. Images are depicted with anterior to the left and posterior to the right. See Techniques and Resources article by Revaitis et al. (dev183210).
Blood-brain barrier disruption in mice lacking FGFBP1 in endothelial cells. Brain blood vessels lacking FGFBP1 (podocalyxin; magenta) fail to deposit collagen IV (green) in the basement membrane and become permeable to circulating compounds (cadaverin; red). See Research article by Cottarelli et al. (dev185140).
The stereocilia bundles are apical extensions of the sensory cells of the inner ear (hair cells), here labelled with phalloidin in a newborn mouse utricle. The development and survival of the hair cells requires the transcription factor GFI1, which functions to repress a neuronal-associated gene expression profile. See Research report by Matern et al. (dev186015).
Interactive 3D model of an E18.5 typically developing mouse skull based on microCT data. Different colours indicate distinct bones. See Research article by Samuels et al. (dev191213).
An eight-cell embryo cultured within the confines of a cylindrical channel in a biocompatible hydrogel, in order to alter the shape and position of its blastomeres, and test the influence of these parameters on cell specification (magenta, pERM; yellow, F-actin; grey, DAPI). See Research article by Royer et al. (dev189449).
A cross-section of the small intestine in an embryonic day 14.5 Ryk knockout mouse (magenta, F-actin; cyan, nuclei). See Research article by Wang et al. (dev195388).
Dorsal view of the nervous system of a Xenopus tropicalis tadpole following β-tubulin antibody staining and confocal microscopy. X. tropicalis is a valuable model for understanding the function of human neurodevelopmental risk disorder genes due to its conserved diploid genome, the ability to make unilateral mutants, and the wealth of experimental tools and knowledge. See Research report by Willsey et al. (dev189290).
ChIP-seq binding data for HOXC6 (left), HOXC9 (center) and HOXC10 (right) in spinal cord patterning. The image was made by Dylan Iannitelli from experimental ChIP-seq data for Hox binding generated by Milica Bulajić. See Research article by Bulajić et al. (dev194761)
Scanning electron micrograph of the junction of the internal and external jugular vein with the superior vena cava in an E16.5 mouse embryo. Venous valves that guard the jugular veins are highlighted in green. Lymphovenous valves that prevent back flow of venous blood into lymphatic sacs are highlighted in magenta. Development of both types of valve is blocked in embryos that lack the RASA1 Ras-GTPase-activating protein, VEGF-C signalling and the transcription co-factors YAP and TAZ. See Research articles by Chen et al. (dev192351) and Cha et al (dev195453).
An image of the moment the optic fissure fuses to create the eye globe in human embryonic development. A section through the eye globe and lens was labelled for the transcription factor PAX6 (blue) and the actin cytoskeleton (white). Cells at the margins rearrange to form two complete epithelial layers. See Research article by Patel et al. (dev193649)
From the 18th to the 20th of November 2020, the “17th Meeting of the Spanish Society for Developmental Biology” (SEBD2020) took place in virtual format, organized by the Spanish Society for Developmental Biology (SEBD), CIC bioGUNE, the University of the Basque Country and the University of Cantabria. The SEBD was founded in 1994 to promote scientific activities in the field of Developmental Biology and to promote the exchange of knowledge within the community of Spanish Developmental Biologists nationally and internationally.
The meeting was initially planned to take place in Bilbao, Spain, on the same dates, but this was not possible due to the COVID-19 pandemic. It is worth mentioning that the first meeting of the SEBD, organized by Juan Aréchaga, took place close to Bilbao in 1996 (University of the Basque Country, Leioa). Little could they imagine the circumstances we would face in the 2020 meeting organization! So, we missed out on the wonderful city of Bilbao, its cultural and food and drink scene, but we did not miss out on the great science being done by developmental biologists in Spain and abroad. Despite the switch to a non-face-to-face format, it was the aim of the organizers to respect the program that was already prepared. We were fortunate that all the invited speakers accepted to change to the virtual format. We still thank them for that!
In the virtual SEBD2020 meeting, different sessions were organized, that ranged from the study of the most basic processes of development (Self-organization, Growth & Scaling, Cell Biology, Evo-Devo, Genomes), to the consequences of the modulation of these processes, such as developmental disease and regenerative biology (Neurodevelopment, Development & Disease, Regeneration). All sessions were initiated by invited speakers, all internationally renowned researchers in Developmental Biology, who presented their unpublished work to an audience of over 250 people ranging from early career researchers to established project leaders. Furthermore, two short presentations and three or four flash-talks were selected per session from the abstracts received.
In the morning of the first day, a virtual outreach event with schools was organized to talk about research and developmental biology with young students all across Spain. After that we had two workshops. The first, on animal experimentation legislation was organized by Augusto Escalante and gave participants tips and tricks on how to build an evaluation report for ethics committees. The second, organized by Sofia J. Araújo and Teresa Rayon brought together a panel to discuss new initiatives on journal-independent peer review. The panel, composed of representatives from innovative projects such as Preprint Review from eLife, Sciety, Peer Community In (PCI), Review Commons and PREreview as well as long standing representatives in preprint curation such as ASAPbio and PreLights from the Company of Biologists, lead a dynamic discussion on the possibilities of changing the peer review system. Concerns were raised about the validity of the review without the label of the signing publisher, although it was also argued that the same concerns can be raised in the case of established publishers. The workshop ended with the overall feeling that it is worth continuing the discussion on which projects will be more valuable for the community.
The Journal independent peer review workshop panelists and some of the participants during the discussion
The scientific meeting kicked off with a plenary seminar by Maria Leptin (EMBL Heidelberg and EMBO Director), on the recent work of her lab on how cell mechanics can modulate intrinsic genetic programmes. This was followed by a session on Growth and Scaling, sponsored by Developmental Dynamics with Marian Ros (IBBTEC, UNICAN) as an invited speaker. Marian presented recently published work from her lab on how Hoxc genes are activated in a colinear manner in the embryonic limb ectoderm and are subsequently transcribed in developing nails and hairs, modulating their growth and scaling. The second session focused on multicellular self-organization, and started with an invited seminar by Miki Ebisuya (EMBL Barcelona) on recently published work from her group showing an elegant example of how mouse ES cells and human iPS cells can autonomously generate oscillating expression patterns that are faster in mouse than in human cells, reflecting the species specific somitogenesis clock period observed in the embryo.
The second day started with the Neurodevelopment session, with an invited talk by Elisa Martí (IBMB-CSIC, Barcelona) who told us how the caudal part of the spinal cord is generated through secondary neurulation, dependent on cell intercalation driven by SMAD3 and YAP. This gave way to the Genomes session, dedicated to the memory of José Luis Gómez-Skarmeta, who recently left us prematurely, leaving a great sadness in all the community. The session was opened by Tatjana Sauka-Spengler (Oxford University, Radcliffe Dept. of Medicine) who focused on the regulation of FoxD3, a gene encoding a key transcription factor during neural crest development. By studying different FoxD3 cis-regulatory elements and using single-cell approaches in zebrafish, Tatjana discussed the role of FoxD3 in controlling the development of trunk neural crest cells from neuromesodermal progenitors. Furthermore, the SEBD organized a tribute video to José Luis, with testimonies from his closest collaborators and friends, that praised the life and scientific achievements of this extraordinary researcher and captured the essence of what the figure of José Luis represents for us. The video was shared with those outside the meeting by YouTube live.
Some of the invited Speakers at the beginning of their presentations.
The afternoon brought us the Cell Biology session, sponsored by the journal Mechanisms of Development, and the talk of Mónica Bettencourt-Dias (Gulbenkian Science Institute Director) on the de novo origin of centrioles, the diversity of centriole structure between species and even within the same organism, and centriole abnormalities in cancer. Later, we had the Development and disease session, sponsored by the journal Developmental Biology. The session opened with an inspiring talk by Ramón Muñoz-Chápuli (University of Málaga), who introduced the audience to the origin and role of endothelial cells in mammals.
The third and last day started with a session in Evo-Devo. The invited speaker Naoki Irie (University of Tokyo, Japan) spoke about the limits that the genetic programs of development impose to the evolution of vertebrates. With an astounding collection of embryos from a plethora of chordate species, Irie presented a comparison of transcriptomes and epigenetic signatures throughout embryonic development and vertebrate species. Last but not least, we had the Regeneration session, representing a growing branch of developmental biology. It started with a seminar by Florenci Serras (University of Barcelona), who shared with us his group results on the mechanisms of sensing and repairing damage in epithelial tissues, using Drosophila wing discs as the model system.
At the end of the meeting, we organized an awards session, where several prizes were announced. The President of the SEBD, Miguel Torres (CNIC), announced the two winners of the “José Luis Gómez-Skarmeta Award to Scientific excellence in Developmental Biology” for young PIs. Alvaro Rada-Iglesias (IBBTEC, UNICAN) and Manuel Irimia (CRG, Barcelona) won this prize ex-aequo for their contributions to genome regulation in development. Also, the “SEBD Awards for scientific excellence in doctoral theses” were announced, and given this year to Diana García Morales (CABD), Pedro Javier Gómez Gálvez (IBiS) and Cristina Sánchez Fernández (UNICAN) for their outstanding Ph.D. work.
Furthermore, during the meeting, there were two science-competitions. The SEBD prizes for the best posters were selected by a scientific committee present at the meeting. The best poster prize went to Carlos Camilleri-Robles from the University of Barcelona, who presented his results on Regeneration of Drosophila wing imaginal discs. The 2nd prize went to Daniel del Toro also from the University of Barcelona for his poster on migrating neuron guidance. And we had two 3rd prizes for Cruz Morenilla-Palao from the Neurosciences Institute and José Santos-Pereira from the CABD-CSIC, on the formation of bilateral circuits and zebrafish gene regulation, respectively.
The three prize-winning images
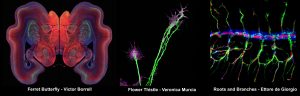
A Photography Competition was sponsored by the journal Developmental Biology. In this SciArt competition there were many beautiful images from different developmental biology models and methods. Victor Borrell (Neurosciences Institute, Alicante) authored the winning image, a ferret kid brain section named “Ferret Butterfly”. The second prize went to Veronica Murcia from the same institute, with a growth cone image she entitled “Flower Thistle”. And, last but not least, Ettore de Giorgio (IBMB-CSIC, Barcelona) got the third prize with a photo of a Drosophila embryonic trachea entitled “Roots and Branches”. There were many other beautiful pictures, which unfortunately could not be selected, but we have been posting them in the SEBD Twitter and Instagram accounts.
Notwithstanding the virtual format, we decided to pursue initial plans for poster presentations and side-activities, aiming to promote as much interaction as possible among participants. Posters were visible in pdf format at the virtual platform and, in many cases, were accompanied by explanatory audio or video. Most of the presentations, were live and followed by a live Q&A session by the audience. This made the sessions highly interactive, dynamic and participative, with lively discussions. Oral presentations were recorded and made available for up to one week after the meeting finished. Discussions in the poster sessions were done through the Slack channels or (virtual) face-to-face in Remo.
Virtual meetings lack human contact and networking which is so important for science advancement. An extra effort from the organizing and scientific committees was done to facilitate computer-based communication tools, including online platforms for live presentations and discussions, Slack channels for specific discussion on each poster and scientific talk, and Remo sessions for informal discussions among participants. Slack channels were also used for putting adds and general information about the meeting. All together these channels helped fill in the gap left by the lack of real-life contacts.
We were very fortunate to count on sponsorship from the Company of Biologists, Developmental Biology, Mechanisms of Development, Developmental Dynamics and Development. This financial support was used to waive the registration fee for undergraduate and PhD students, which benefitted students not only form the SEBD, but also from abroad (other European countries, and other as far as Brazil, Mexico and Australia). Opinion polls were performed at the end of the meeting, where most of the participants manifested that the meeting was better or much better than they anticipated! And nobody complained about the catering…
The SEBD2020 Scientific Committee: Isabel Almudi, Sofia J. Araújo, Rosa Barrio, Laura Bozal, Augusto Escalante, Fernando Garcia-Moreno, Maria Losada-Perez, Ignacio Maeso, Luciano Marcon, Oscar Ocaña, Olatz Pampliega
The enteric nervous system (ENS) derives from the neural crest and innervates the gastrointestinal system, in which it is essential for gut function throughout life. A new paper in Development uses zebrafish to investigate the poorly understood process of post-embryonic ENS neurogenesis, in both development and injury contexts. To find out more, we met the paper’s two authors, Wael Noor El-Nachef, Assistant Clinical Professor of Medicine at UCLA, and Marianne Bronner, Albert Billings Ruddock Professor of Biology and Biological Engineering at Caltech.
Wael (L) and Marianne (R)
Marianne, can you give us your scientific biography and the questions your lab is trying to answer?
MB: When I started graduate school, I thought that I wanted to do structural biology, but then I took a course in developmental biology and found my passion. I learned about Nicole Le Douarin’s work on the neural crest and it blew me away. I immediately knew that I wanted to work on neural crest cells and be just like her. To this day, she remains my role model. I was fortunate enough to get my first faculty position at the University of California Irvine right out of graduate school. I then moved to Caltech in 1996, where my lab grew larger but continued working on the same types of problems, while adapting new and exciting technologies to help solve them.
My lab pursues diverse questions in neural crest biology, ranging from how and when neural crest cells are induced to how they may have arisen and changed during vertebrate evolution. How and why have neural crest cells continued to acquire new traits? How do they migrate, find their proper sites and differentiate into a diverse array of cell types? Most recently, we have become fascinated by their potential role in tissue regeneration. Wael’s work on de novo neurogenesis in the ENS fits perfectly into these areas of interest.
And Wael – how did you come to work in Marianne’s lab and what drives your research today?
WE-N: I have had somewhat of a non-traditional path into basic science research in that I didn’t get serious about research until after medical school. During my clinical training, I was frequently frustrated by so many ‘idiopathic’ diseases in gastroenterology that are difficult to treat and terrible for the patient, and the most challenging of those conditions often involve the ENS. Rather than wait around for someone else to figure out these problems, I decided to pursue research training in regenerative medicine, initially studying cultured enteric neuronal progenitors in a tissue-engineered intestine construct. Although that was a rewarding experience, I quickly realized that there are many fundamental issues that we do not yet understand about ENS development and homeostasis. And without establishing some of these basics, regenerative medicine approaches to enteric neuropathies will be stymied.
My big revelation came when I realized that ‘developmental biology’ is not limited to embryogenesis, but rather includes postnatal development and growth, aging and senescence, and homeostasis and regeneration. Or, in the words of John Wallingford, ‘we are all developmental biologists’. This perspective expanded my views of the field I wished to join and I reached out to Marianne (I cold called her, and she surprisingly took me on). Training in her lab seems like an obvious choice in retrospect, but in our sometimes cloistered research niches it initially felt a bit mavericky. Luckily, UCLA, my home institution, has an established affiliation with Caltech and has been extremely supportive of my research pursuits.
How has your research been affected by the COVID-19 pandemic?
WE-N: Like so many other scientists, our experimental lab work was placed on hold for a few months while we were on lockdown. Luckily, we are currently ramping-up our activity in the lab; we work in shifts, practice physical distancing, wear masks, etc. It’s great to be back in the lab, and I’m sure I’m not the only one who may have gotten a bit misty-eyed pipetting something for the first time in months. I consider myself extremely fortunate in that I didn’t lose anything other than some time; my family and I are healthy, my fish lines are intact, my projects can restart fairly easily, etc. But returning to the lab within this current COVID-19 paradigm, I now realize the importance of those chance collisions with my colleagues when we informally exchange ideas or troubleshoot a problem together during our daily lab tea time. Videoconferencing is no substitute.
Why has the manner (and even the existence) of postnatal enteric neurogenesis been so controversial?
WE-N: It comes down to this: the gastrointestinal tract is difficult to study. It is arguably the largest endocrinologic organ, largest epithelial organ, largest immunologic organ and second largest neurologic organ; it fills up the largest body cavity in a long convoluted bundle; it is constantly moving and is dynamic in several respects; and to top it all off, it is filled with ‘luminal contents’, which include the largest microbiome compartment of our body. Although this amazing complexity is what first attracted me to this field, there are days when I envy those who study the central nervous system.
In model organisms other than zebrafish, one challenge has been how to directly visualize and detect enteric neurogenesis. Simply counting neurons from a representative section and then extrapolating in some way is not reliable. Live imaging the ENS in species such as mouse is possible but technically challenging and often carried out using a few time points rather than a several-hours long time-lapse. Lineage-tracing experiments with murine Cre transgenic lines have been hampered by relatively low recombination rates. Assessing for evidence of enteric neurogenesis with the use of thymidine analogues has also been complicated by unclear optimal pulse/chase duration and timing. Lastly, studies of ENS homeostasis using non-zebrafish models often investigated the intestinal tract divorced from the rest of the body, and thus may have been unable to consider the possibility of a gut-extrinsic source of neuronal precursors using their methods.
Ultimately, new enteric neurons may arise in small numbers over time and over a large area, and thus may be difficult to detect without fully surveying the ENS. It is also important to acknowledge that alternative hypotheses are not necessarily mutually exclusive, and nature may have provided multiple solutions for addressing issues of ENS homeostasis. But, my experience with the zebrafish model has shown me the advantages of using a model that is highly tractable and amenable to live and in toto imaging to allow us to compellingly explore questions of enteric neurogenesis.
Can you give us the key results of the paper in a paragraph?
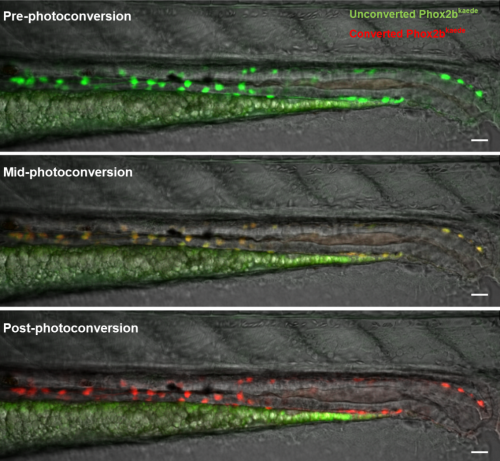
WE-N & MB: Using a photoconvertible transgenic line to ‘time stamp’ enteric neurons, we found that enteric neurogenesis persists in post-embryonic zebrafish development, despite an apparent absence of enteric neuronal progenitors, including enteric glia. Lineage tracing with carefully timed DiI injections into the neural tube, as well as with an inducible Cre-transgenic line, supported a trunk neural crest origin of these post-embryonic enteric neurons, consistent with them arising from Schwann cell precursors (SCPs). Enteric neurogenesis was also observed after injury, modelled with two-photon laser ablation of individual neurons. The 5-HT4 receptor agonist prucalopride increased enteric neurogenesis during normal development, as well as in pre-treated fish that subsequently underwent neuronal ablation.
Images of a 4.5 days postfertilization zebrafish midgut and hindgut undergoing photoconversion (enteric neurons are green at baseline, gradually become orange and ultimately red).
Comparing zebrafish with other species, what can you infer about the contribution of SCPs to the ENS during evolution?
WE-N & MB: In the lamprey, an early jawless vertebrate that appears to lack a vagal neural crest, the ENS originates from the trunk neural crest, suggesting that this may have been the original strategy for providing the basal vertebrate intestine with enteric neurons. However, the evolution of jawed vertebrates was accompanied by the appearance of the vagal neural crest, and the initial formation of the ENS in these animals arose from this newer cell population. We suspect that rather than completely losing the trunk crest contribution to the ENS, later vertebrates adapted this cell population of SCPs to act as a reservoir of ENS progenitor cells that contribute enteric neurons as the intestine grows or to maintain enteric neuron number in the case of injury.
Does your work have any implications for the understanding or treatment of human enteric neuropathies?
WE-N & MB: Certainly. As there is evidence of SCP-derived enteric neurogenesis in teleosts, chicks and mice, it is reasonable to consider that this is also conserved in humans. Many acquired enteric neuropathies are thought to be due (at least in part) to a loss of enteric neurons. In such disease states, promoting enteric neurogenesis with agents such as 5HT4 receptor agonists represents a potential therapeutic approach that is much simpler than trying to rebuild the ENS by injecting precursors that were created in a lab.
Stepping aside from treatment, understanding why a patient’s SCPs are unable to maintain enteric neuron number may lead to a deeper understanding of these pathologies. Is the enteric neuron loss too profound for the SCPs to compensate, or does the disease process involve the SCPs directly? Could enteric neuropathies actually be SCP-opathies?
When doing the research, did you have any particular result or eureka moment that has stuck with you?
WE-N: My first time-lapses that captured new enteric neurons arising in the intestine really energized my efforts. To me, it was exciting, aesthetically pleasing to view, and, simply put, cool. That feeling of seeing something that perhaps no one else has observed before is a very special feeling. I did those experiments early on, and it gave me the confidence that I had the right tools and techniques to explore my research questions in fun and interesting ways.
Seeing something that perhaps no one else has observed before is a very special feeling
And what about the flipside: any moments of frustration or despair?
WE-N: I love coffee. But it turns out that too much coffee on the day you need to dechorionate and microinject zebrafish embryos is not a good thing. Suffice to say that day of experiments was an absolute disaster and had to be rescheduled.
What’s next for you after this paper?
WE-N: I’m very interested in exploring the evolutionary development of enteric glia further. Our results suggest that zebrafish may be missing enteric glia, or at least cells with classical glial properties in the gut. If we can determine when in evolution enteric glia arose, it may help us infer why they arose. Currently, there is some controversy concerning the role of enteric glia in health and disease, and improving our understanding of their function may shine a light on a number of gastrointestinal diseases.
I’m also looking forward to better defining the SCP cell population in zebrafish to aid me in the study of their migration dynamics, and to begin exploring the potential signalling mechanism of 5HT4 receptor agonism in enteric neurogenesis.
Where will this story take the Bronner lab?
MB: We are very interested in evolutionary changes in the ENS and how and when increased complexity arises. Comparing diverse species, such as lamprey, zebrafish and mouse ENS neurons and glia, may offer clues as to how the ENS has become elaborated during the course of vertebrate evolution.
Another direction we are pursuing is using single cell RNA sequencing to examine the heterogeneity of enteric neurons in zebrafish as a function of time. Despite the fact that the zebrafish intestine is comparative simple compared with mouse, there are a diverse array of neurons characterized by different neurotransmitters and neuropeptides. Understanding the molecular underpinnings of how and when these differentiate fascinates me and is a direction we are currently pursuing.
Finally, let’s move outside the lab – what do you like to do in your spare time in California?
WE-N: I have a 1-year-old son, and I’ve recently taken up hiking with him sitting in a special pack that I wear on my back. If I slow down, he’ll bang on my head, so it’s a great workout with very personalized coaching.
MB: I enjoy swimming and hiking, both of which I have been lucky enough to continue during the lockdown. My baking has also improved!
 (No Ratings Yet)
(No Ratings Yet)


 (5 votes)
(5 votes) Understanding brain circuit evolution at single-cell resolution using comparative connectomics and transcriptomics
Understanding brain circuit evolution at single-cell resolution using comparative connectomics and transcriptomics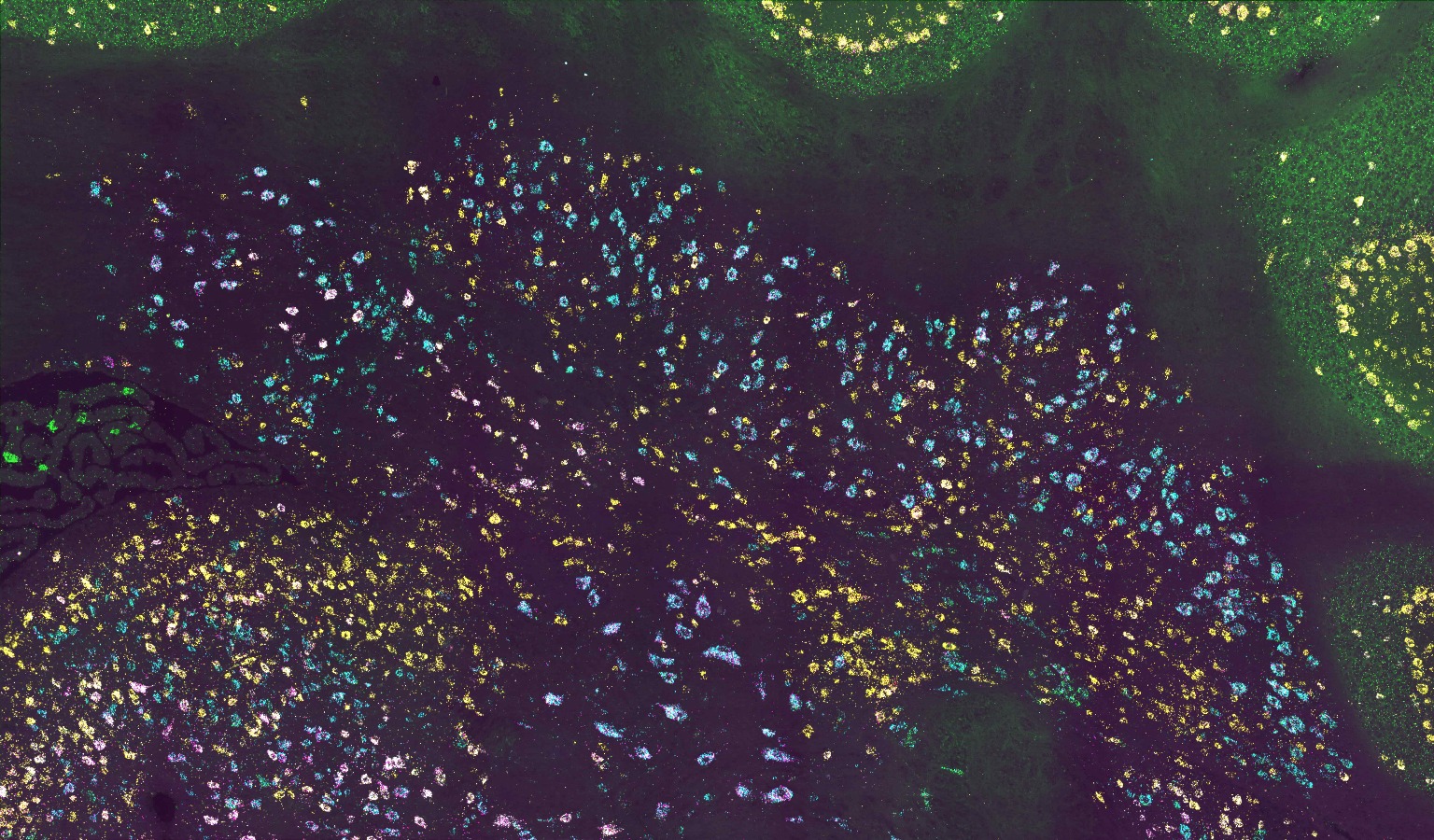
![Micrographs of C. elegans anchor cells (cdh-3>mCherry::moeABD, magenta) atop the uterine-vulval basement membrane (laminin::GFP, green), expressing endogenously-tagged transcription factors (nuclear, green), grouped by column (left to right: egl-43::GFP, fos-1::GFP, hlh-2::GFP and nhr-67::GFP). Each row represents a different RNAi perturbation [top to bottom: empty vector, egl-43(RNAi), fos-1(RNAi), hlh-2(RNAi) and nhr-67(RNAi)]. See Research article by Medwig-Kinney et al. (dev185850)](https://i0.wp.com/thenode.biologists.com/wp-content/uploads/2020/12/develop_147_01_cover-scaled.jpg?w=121&h=158&ssl=1 "JEBcover_Front")
























 In the morning of the first day, a virtual outreach event with schools was organized to talk about research and developmental biology with young students all across Spain. After that we had two workshops. The first, on animal experimentation legislation was organized by Augusto Escalante and gave participants tips and tricks on how to build an evaluation report for ethics committees. The second, organized by Sofia J. Araújo and Teresa Rayon brought together a panel to discuss new initiatives on journal-independent peer review. The panel, composed of representatives from innovative projects such as
In the morning of the first day, a virtual outreach event with schools was organized to talk about research and developmental biology with young students all across Spain. After that we had two workshops. The first, on animal experimentation legislation was organized by Augusto Escalante and gave participants tips and tricks on how to build an evaluation report for ethics committees. The second, organized by Sofia J. Araújo and Teresa Rayon brought together a panel to discuss new initiatives on journal-independent peer review. The panel, composed of representatives from innovative projects such as