With this strange and difficult year drawing to a close, I wanted to take the opportunity to update our community on how things have been going at Development through pandemic times. In March, the entire staff of The Company of Biologists decamped from the office and started home working. We’re incredibly lucky to have secure jobs that can be done efficiently from home, and while there have certainly been some challenges with adapting to remote working, things have gone surprisingly smoothly and we have (we hope!) continued to provide a high level of service to our authors and readers.
We have been monitoring our activities and workflows closely since COVID-19 hit, and I have to say that, if you looked just at these metrics, you would hardly know the extent of the disruption we’ve all experienced this year. As labs across the world shut down, we thought we might either see a flood of submissions as researchers had more time to write or – conversely – a significant fall as people had so many other challenges to deal with and/or were unable to finish key experiments. What actually happened was neither of these things: the rate of both initial submissions and revisions has essentially kept pace with previous years.
We thought we might receive more short ‘Report’ format articles as projects were written up earlier than they might have been otherwise, but this has not been the case. For the same reason, we wondered whether there might be a dip in the quality of papers submitted, but editorial rejection and overall acceptance rates have held steady. One thing we have seen is a slight shift in the geographic origin of papers submitted to us, with more papers from China-based authors and fewer from Europe. Whether this is a consequence of the pandemic or a more general trend, though, is hard to tell at this point – particularly given that submissions from China have been growing in recent years anyway.
Something we definitely expected was a slowdown in our decision speeds, especially in the early months. We knew that our editors and reviewers were facing the challenges of closing down their labs and/or making them COVID-safe, moving their teaching and mentoring online and – in many cases – taking on additional caring and home-schooling responsibilities. And we did find that some papers were significantly delayed through editorial assessment and peer review; for those authors that did experience such delays, we’re sorry. But, amazingly, our average turnaround times have remained the same this year as last. And even more impressively, more referees accepted our invitations to review papers, and more of them returned their reports on or before their deadline.
One issue that particularly concerned us was the potential disproportionate effect of the pandemic on women. Several reports earlier in the year suggested that submission rates from women were likely to decrease as they bore the brunt of childcare (see e.g. https://www.nature.com/articles/d41586-020-01294-9). So have we seen any evidence of this at Development? We don’t record author or reviewer gender in our submission system (though this is something we’re working on) so any analysis is necessarily imperfect. But by running names through the genderize.io app, we can get some idea of the gender demographics of our community (I’d note, though, that genderize.io is particularly bad at accurately assigning gender to non-Western names, and of course that this gender assignment is binary and therefore inherently limited). In 2018/2019, 31% of corresponding authors and 48% of first authors for whom gender could confidently be assigned were women. In 2020 (to the end of October), those numbers were 32% and 49%. What about referees? Again looking only at those individuals for whom we could confidently assign gender, 35% were women as compared to 32% in 2018/2019. Pre-pandemic, women were slightly less likely to accept an invitation to review than men, but this year, that trend has reversed. Overall, the data suggest an increasing representation of women among both our author and reviewer pool in recent years, and we hope this trend will continue.
The fact that the metrics for 2020 have looked so ‘normal’ does not mean that researchers have not found the year incredibly tough. Rather, it pays testament to the dedication and resolve of our community to keep going through these difficult times. And while 2021 brings the promise (fingers crossed!) of widely-accessible vaccines, the pandemic is not yet over, and the inevitable economic downturn will present its own financial challenges to the research sector. So I’d like to thank everyone who has contributed to and interacted with the journal this year, and who will do so in the year(s) to come – your support is hugely appreciated.
Unregulated cell proliferation can be disastrous for development and underlies the progression of cancers throughout the lifespan. A new paper in Development dissects the molecular regulation of a key cell proliferation promoter (and infamous oncogene) Myc, using Drosophila as a model system. We caught up with Olga Zaytseva, recent PhD graduate and one of the paper’s first authors, and her supervisor Leonie Quinn, Associate Professor at the John Curtin School of Medical Research in Canberra, to find out more.
Olga (L) and Leonie (R)
Leonie, can you give us your scientific biography and the questions your lab is trying to answer?
LQ After my PhD studies in Adelaide I conducted postdoctoral research at the Peter MacCallum Cancer Centre, investigating growth and cell cycle control. In 2007, I established my own group at the University of Melbourne, and later relocated to the John Curtin School of Medical Research in Canberra. My laboratory uses Drosophila to elucidate molecular mechanisms patterning cell and tissue growth during animal development. Our interest in RNA-binding proteins ‘moonlighting’ in transcriptional roles began over a decade ago when we demonstrated the splicing factor Hfp inhibits cell growth through transcriptional repression of the master regulator of growth, Myc. Subsequent studies revealed that a second splicing factor, Psi, also interacts with RNA Polymerase II machinery, in this case activating Myc transcription to promote cell and tissue growth. In our current paper, we report that the Argonaute family RNA binding protein, AGO1, functions outside of canonical functions in the miRNA-dependent RNA-induced silencing complex (RISC), repressing Myc transcription to inhibit cell and tissue growth.
And Olga – how did you come to work in Leonie’s lab and what drives your research today?
OZ During my undergraduate studies at Melbourne University on a path to a medical degree, I became particularly fascinated by studies of animal development, so I decided to seek out opportunities for research experience in this area. After seeing several beautiful images of the Drosophila wing on the Quinn lab website, I decided to approach Leonie and with her help I was lucky enough to receive a summer studentship to complete a short project on the roles of the transcription factor ASCIZ, previously uncharacterised in flies. I felt very inspired by Leonie’s passion, and this experience of the excitement and rewards of scientific enquiry motivated me to continue on to Honours and then a PhD degree under Leonie’s supervision. My PhD, which I have recently completed, gained insights into how the transcription regulator and single stranded DNA/RNA binding protein Psi acts to control growth and development. From the studies conducted in our lab, I am particularly interested in how transcription regulators such as Psi are able to control particular networks of genes in different cell and tissue types.
How has your research been affected by the COVID-19 pandemic?
OZ & LQ We feel quite fortunate in Australia, given the low number of cases and the stringent measures taken by the government to prevent the spread of infection and to widely test the population. The preventative lockdown measures meant that we’ve had limited access to the lab for several weeks, but given that many lives have been saved as a result of the restrictions, they are entirely warranted. Procedures are already underway to resume operations which means it’s not too long until our experiments can be gradually commenced again.
Let’s get to your current paper then – what was previously known about the link between Myc, Argonaute proteins and growth control?
OZ & LQ Although Myc is a well-established regulator of cell and tissue growth in both mammals and Drosophila, the links between AGO1 and Myc-driven growth have not been investigated. AGO1 proteins have been most thoroughly characterised as having roles in the RISC, where they are guided by the sequences of particular miRNAs/miRs to degrade target mRNAs. The regulation of Myc mRNA via RISC and miR-308 has been previously demonstrated by Julie Goodliffe’s group in Drosophila embryos; however, many miRs have tissue-specific targets and our work has shown that in the context of the wing, AGO1 does not control Myc via a RISC-dependent mechanism. Our study adds to the body of work that implicates Argonaute proteins in transcriptional control within the nucleus, for example via Polycomb Group transcriptional repressors.
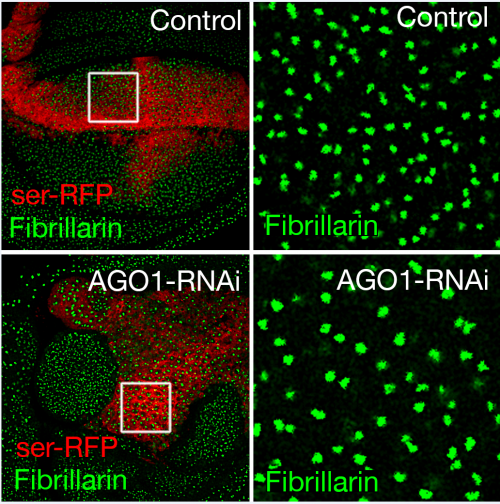
AGO1 knockdown increases nucleolar size.
Can you give us the key results of the paper in a paragraph?
OZ & LQ We have discovered a previously unreported role for AGO1 as a repressor of Myc transcription and, as a result, growth of the Drosophila wing. We depleted AGO1 specifically in the developing wing, and observed strikingly increased ribosome biogenesis, nucleolar size and cell growth. AGO1 physically interacts with a key regulator of Myc transcription, Psi. AGO1 depletion increased Myc mRNA and protein levels, and the overgrowth seen after AGO1 knockdown was dependent on Psi and Myc. Given the roles of AGO1 within RISC, we tested whether Myc was increased via the most likely miR candidates; however we did not observe changes to Myc mRNA after manipulation of miRs. Instead, we saw that AGO1 depletion promotes the RNA Polymerase II transcription of Myc, and that AGO1 directly associates with the Myc promoter.
Do you have clues as to how, at a molecular level, AGO1 inhibits Myc transcription?
OZ & LQ We saw that overgrowth as a result of AGO1 knockdown was dependent on Psi; therefore a likely possibility is that the interaction between AGO1 and the Myc promoter might repress the activity of Psi. The mammalian orthologue of Psi, FUBP1, acts to remodel single-stranded DNA at the MYC promoter which enables maximal RNA Polymerase II transcription. However, links between human AGO1 and FUBP1 have not been studied and would be interesting to follow up in the future. We also saw overlap between AGO1 and Polycomb in the nucleus, and given that previous studies implicated Polycomb in the autorepression of the Myc gene in response to elevated Myc, Polycomb may play a role in AGO1-mediated Myc repression in certain contexts. Furthermore, AGO1 is an RNA-binding protein – the repression of the Myc promoter may occur by recruitment of non-coding RNAs that restrict transcription.
Too much Myc expression is associated with most human cancers – might AGO1 be a useful way to dampen this activity?
OZ & LQ While increased levels of Myc are clearly important for tumour progression, the links between AGO1 and cancer are not well established. Across different tumour types, AGO1 can be either overexpressed or contain loss-of-function mutations, suggesting either oncogenic or tumour suppressive roles depending on the context. The wing epithelium, where we observed tumour suppressive activity of AGO1, provides insights into progression of epithelial tissue cancers such as breast or prostate. A further understanding of how AGO1 is recruited to the Myc promoter would be required to develop strategies that stimulate AGO1 Myc-repressive activity. However, given the potent Myc activation as a result of AGO1 knockdown, harnessing the repressive roles of AGO1 to inhibit cancer growth is an exciting prospect.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
OZ For me, it was seeing the overgrown cells after AGO1 depletion: that was the moment which started the enquiry into how it might occur. Once we began to characterise the phenotype, including elevated ribosomal RNA, ribosomal proteins and increased nucleolar size, we gained more clues that pointed towards a link with Myc. Given the crucial roles Myc plays in many cancers, it’s exciting that we discovered a mechanism which could contribute to cancer progression.
And what about the flipside: any moments of frustration or despair?
OZ When seeking to determine whether AGO1 interacts with chromatin in the Drosophila wing, we initially took a genome-wide approach. However, after making the constructs and performing the sequencing, we didn’t get any enrichment signal, which was quite disappointing. Therefore we looked at the Myc promoter specifically and were able to observe AGO1 binding. However, any clues as to AGO1 transcriptional targets more widely in the cell would have been highly informative, and it’s an experiment we’d like to perform in the future.
What next for you after this paper?
OZ I’m very interested in the protein that led us to AGO1, Psi. Both Psi and the mammalian orthologue FUBP1 appear to play versatile roles in binding RNA and single-stranded DNA at promoters. In fact, there are many examples of proteins that originally were thought to exclusively bind RNA but were subsequently found to associate with DNA as well. This highlights the dynamic nature of DNA, which often doesn’t get considered – the textbook ‘double helix’ is in fact just one of the conformations, while many other arrangements of the DNA strands play significant regulatory roles. Understanding how proteins such as Psi interact with such structures is important for a full picture of transcriptional control. As a first step, we are investigating genome-wide targets of Psi which led us to the discovery that Psi regulates multiple genes with roles in growth and developmental patterning.
There are many examples of proteins that originally were thought to exclusively bind RNA but were subsequently found to associate with DNA as well.
Where will this work take the Quinn lab?
LQ Based on this work we will continue to investigate the nuanced context-dependent control of transcription by single-stranded DNA/RNA-binding proteins, which is a key interest of our lab.
Finally, let’s move outside the lab – what do you like to do in your spare time in Canberra?
LQ I voraciously read non-fiction – history and politics of all flavours.
OZ I enjoy getting outdoors to explore the many parks and forests around Canberra.
Videos of the talks and Q&As can now be found here!
We’re happy to confirm the next in our Development presents… webinar series will be chaired by our Editor Yrjö Helariutta (Sainsbury Laboratory, University of Cambridge / Institute of Biotecnology, University of Helsinki) and features three talks on various aspects of plant development.
Wednesday 2 December 2020 – 10:00 GMT
Marta Mendes (from Lucia Colombo’s lab at the University of Milan)
‘The RNA dependent DNA methylation pathway is required to restrict SPOROCYTELESS/NOZZLE expression to specify a single female germ cell precursor in Arabidopsis’
Krisztina Ötvös (from Eva Benková’s lab at IST Austria)
‘Modulation of root growth by nutrient-defined fine-tuning of polar auxin transport’
Kenji Nagata (from Mitsutomo Abe’s lab at the University of Tokyo)
‘Ceramides mediate positional signals in Arabidopsis thaliana protoderm differentiation’
The webinar will be held in Remo, our browser-based conferencing platform – after the talks you’ll have the chance to meet the speakers and other participants at virtual conference tables. If you can’t make it on the day, talks will be available to watch for a couple of weeks after the event (look out for details on the Node).
For more information about what to expect in Remo, go to
We seek a motivated and organized individual to join the Lyons Lab (https://www.lyonslab.org/) at the Scripps Institution of Oceanography (https://scripps.ucsd.edu/) to serve as lab manger and research assistant. The individual will assist the lab in funded projects on molluscan neurodevelopment, developmental gene regulatory networks, and biomineralization. The successful candidate will be responsible for lab organization, inventories, safety protocols, and care of colonies of marine invertebrates. In addition, the individual will conduct lab research such as PCR/cloning, in situ hybridization, microinjection/electroporation, collection of embryonic samples for RNAseq/ATAC-seq/scRNAseq, and microscopy. Previous experience with the above techniques, particularly research with embryos of marine invertebrates, is preferred but not required. The successful candidate will have exceptional organizational and record-keeping skills and will serve as a collaborator with other lab members. The Lyons Lab is dedicated to building and maintaining a diverse and inclusive lab culture. The Scripps Institution of Oceanography, a department at U.C. San Diego, provides an exceptional learning and research environment within the large San Diego Metropolitan Area. The application closes December 9th, and the position will be filled as soon as possible.
Our laboratory at the Federal Institute of Technology (EPFL) in Lausanne, Switzerland is looking for a postdoc to work in the field of gene regulation during mammalian development, by using in vitro cultured, ES cells-derived gastruloids (pseudo-embryos) as a model system. Key-words: Transcriptional regulation, enhancers, CRISPR/cas9, mutant gastruloids, epigenetic profiling, 3D chromatin structure, single-cell RNA-seq phenotyping, DORA, open science, Hox genes (but not only). We look for an independent and respectful colleague, with a strong interest and curiosity for basic research and life in general. Fully funded position.
Disease Models & Mechanisms, published by The Company of Biologists, is seeking enthusiastic and motivated applicants for the role of Features & Reviews Editor. Joining an experienced and successful team, including Editor-in-Chief Liz Patton, this is an exciting opportunity to make a significant contribution to a growing Open Access journal in the fast-moving field of translational research.
This is a permanent, full-time position, and is based in The Company of Biologists’ attractive modern offices on the outskirts of Cambridge, UK. Owing to the pandemic, an initial period of remote working may be necessary. Please see the full job description for further details.
To apply, or for more information, contact recruitment@biologists.com. Applications should be made as soon as possible and by 21 December 2020. Applicants should be eligible to work in the UK.
There is a 4-year PhD position available at the Sars International Centre for Marine Molecular Biology (www.sars.no/) in the lab of Dr. Patrick Steinmetz.
The successful applicant will study how starvation-induced degrowth is controlled in the sea anemone Nematostella vectensis. The project may also involve studying key aspects of degrowth in the sea anemone Aiptasia pallida in a comparative context. State-of-the-art functional techniques (CRISPR mutants & knock-in lines, transgenesis), and a diverse range of approaches from developmental biology, genetics, physiology and genomics are available for functional and genomic studies in Nematostella.
Further details and how to apply: https://tinyurl.com/y3h5tv8r
IMBA is opening a search for a group leader. It is an absolutely great place to do 100% basic research. Full freedom, no questions asked. You can find all info here. The deadline is the 10th of January.
Dawei Sun has just finished his PhD in Dr Emma Rawlins’ lab at The Gurdon Institute here in Cambridge, UK. He has remained in the city with his wife for the duration of the pandemic. In the middle of the UK’s second lockdown we met him, virtually, to find out how he is doing, hear about the triumphs and tribulations of his PhD, and discuss his experience of studying in the UK. (more…)
Nuclear-encoded mitochondrial ribosomal proteins (MRPs) are crucial for mitochondrial function and hence cellular energy production. A new paper in Development investigates the function of MRPs in early mouse development, thanks to a battery of knockout mutants. We caught up with first author (and recent graduate) Agnes Cheong and her supervisor Jesse Mager, Associate Professor at the Department of Veterinary and Animal Sciences at the University of Massachusetts, Amherst, to find out more.
Agnes and Jesse at Agnes’ PhD defence – note the mouse cake
Jesse, can you give us your scientific biography and the questions your lab is trying to answer?
JM I did my PhD work with Terry Magnuson at UNC (University of North Carolina at Chapel Hill), studying the Polycomb Group Gene EED and connecting PRC2 with genomic imprinting, X-inactivation and gastrulation defects. I then did postdoctoral work with Rudolf Jaenisch and Marisa Bartolomei, again investigating epigenetic regulation during early development, which I have continued in my own group since 2008 at UMass. We continue to identify novel parent of origin events during gastrulation, as well as characterizing a large number of early lethal phenotypes arising from the Knockout Mouse Project that advance our understanding of the diverse array of genes required for implantation and gastrulation.
Agnes, how did you come to work in Jesse’s lab and what drives your research today?
AC My first research experience involved the study of mammalian implantation and I was really fascinated by the remarkable transformations the embryos undergo during early development. I wanted to continue exploring my interest in embryogenesis, which was the reason why I decided to stay at Jesse’s lab after my rotation. We know so much more than we did decades ago, yet when we take a magnifying glass to zoom into different developmental processes, many blank spaces remain. The desire to comprehend this convoluted process is my motivation to pursue research in development.
When we take a magnifying glass to zoom into different developmental processes, many blank spaces remain
How are each of you coping in the current COVID-19 pandemic?
AC A workday at home is certainly very different from that at the lab. Since I started working from home, I lost the privilege of casually reaching my 10,000 steps daily. Besides my baking hobby, I am otherwise eager to be able to get back to the lab and do some bench work. Fortunately, the Mager lab still has regular virtual lab meetings so that we can present our research ideas and catch up with our lab family, even though we are not able to be physically present together.
JM Since the shutdown mid-March, the lab is now basically just me and the mice…so a bit lonely. But we’ve had lots of Zoom meetings and are trying to keep the lab members busy writing and reading. It definitely took a few weeks to find productive work patterns (and bonus regular lunches with my kids!). Daily evening strolls with my family have been a silver lining – we are fortunate to be able to get outside quite a bit.
Let’s get to your current paper then, which uses mutants produced by the Knockout Mouse Project. What were the aims of that project and your lab’s involvement with it?
AC & JM IMPC/KOMP is a global initiative, and its main goal is to generate a comprehensive and public resource comprising mouse embryonic stem cells and mice containing a null mutation in every gene within the mouse genome. Although the bulk of the effort has focused on adult phenotypes, ∼25% of nulls are lethal and 12% die prior to E9.5. Our involvement with KOMP is to characterize a large number of these early lethal phenotypes since there is no planned pipeline to analyse early embryos. Thus far, we have phenotyped more than 110 gene knockouts, and will grow to ∼250 over the next few years. Check out all our results at http://blogs.umass.edu/jmager/.
Can you give us the key results of the paper in a paragraph?
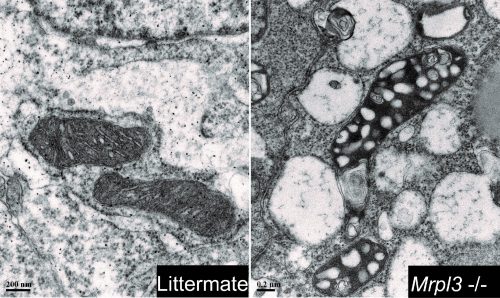
AC & JM Our paper demonstrates the importance of a functional oxidative phosphorylation (OXPHOS) system during early development. Although there are over 70 mitochondrial ribosomal protein (Mrp) genes in mammals, the absence of a single Mrp gene results in pre-gastrulation developmental arrest. Despite the presence of pluripotent epiblast cells in the Mrp mutants, these embryos fail to initiate EMT and progress through gastrulation. This disruption is the result of a malfunctioning mitochondrial translation system which contributes to a lack of ATP production, and hence results in cell cycle disruption and stalling at the G2/M checkpoint. Our results are aligned with other developmental studies of mitochondrial functioning genes showing that the OXPHOS is indispensable during early embryogenesis.
TEM images revealing the different mitochondrial morphologies between mutants and control embryos.
Before developmental arrest, early embryonic development runs normally in Mrp mutants – does this mean the MRPs are doing nothing before then?
AC & JM Not necessarily: since the majority of Mrp genes are also present in the oocyte, maternal Mrp protein could be supporting development through implantation and/or an alternative energy system, such as glycolysis, may be a major factor driving developmental progression prior to gastrulation. A more-detailed study of protein turnover and maternal deletion experiments will provide insights on the precise time point at which Mrp proteins are essential during embryogenesis.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
AC My memorable eureka moment was when I analysed phospho-cdc25c foci and found a significant increase in the mutants, confirming my suspicion that the malfunctioning OXPHOS system leads to cell cycle disruption.
And what about the flipside: any moments of frustration or despair?
The endless sea of genotyping when working with 20+ different knockout lines and needing to genotype every mouse and embryo.
So what next for you after this paper?
AC I recently completed my PhD defence. My plan is to diversify my scientific experience and continue my research career in a biotech or industrial setting.
Where will this work take the Mager lab?
JM Our findings that so many nuclear-encoded mitochondrial functioning genes have such similar phenotypes (gastrulation arrest) are prompting the design of experiments to try to assess the metabolic needs of distinct pre-gastrulation lineages and cell populations. This is a new and exciting area or inquiry for our group. We will also pursue conditional knockout approaches to examine the maternal protein contribution.
Finally, let’s move outside the lab – what do you like to do in your spare time in Amherst?
AC Amherst is a small college town surrounded by many local breweries. When the weather is nice and warm, I enjoy walking to town and reward myself with a hoppy and cold beer. On cold rainy days, I like warming up my apartment with baked goods and sharing them with my friends.
JM I’ll be honest – I’m an exercise enthusiast (to put it mildly)…so if I’m not at work or with my family I’ll generally be found getting those endorphins.
 (2 votes)
(2 votes)
 (No Ratings Yet)
(No Ratings Yet)
 There is a 4-year PhD position available at the Sars International Centre for Marine Molecular Biology (
There is a 4-year PhD position available at the Sars International Centre for Marine Molecular Biology (