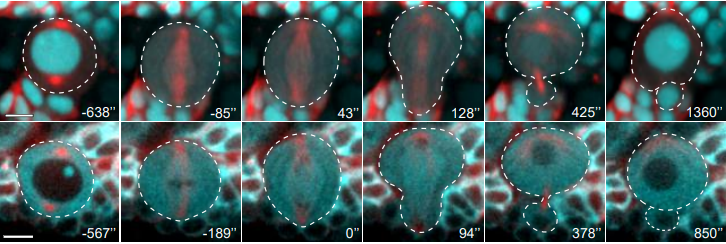
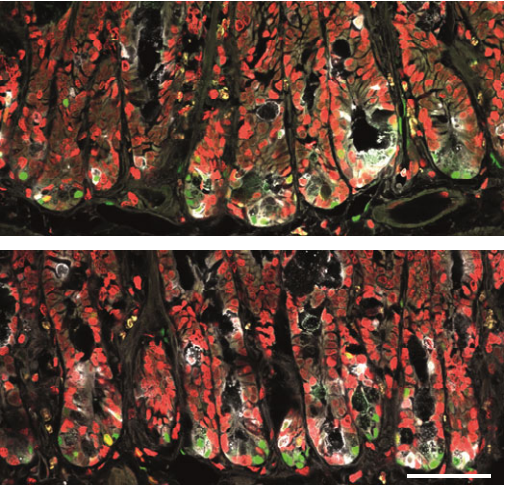
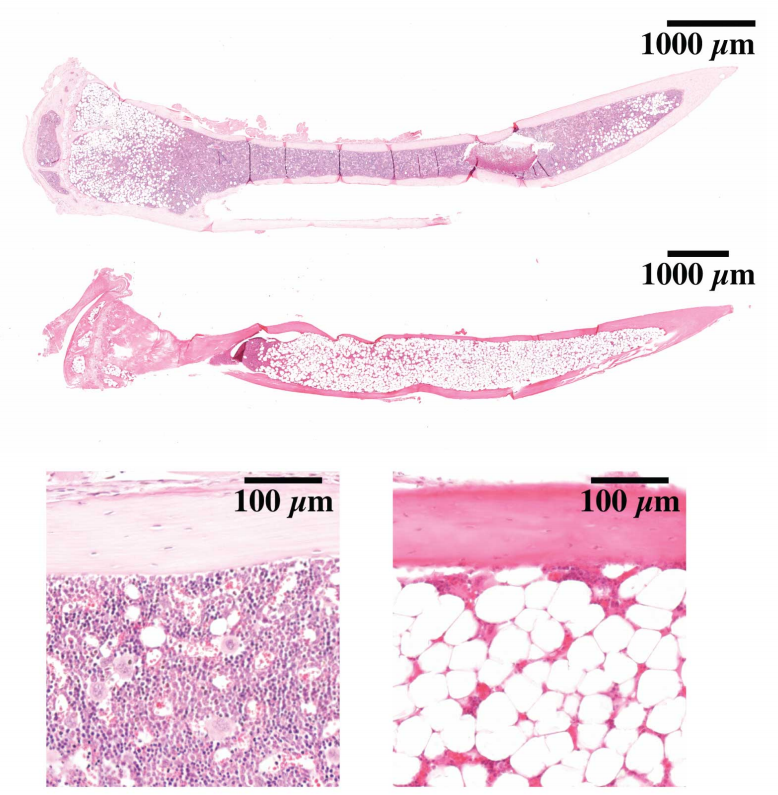
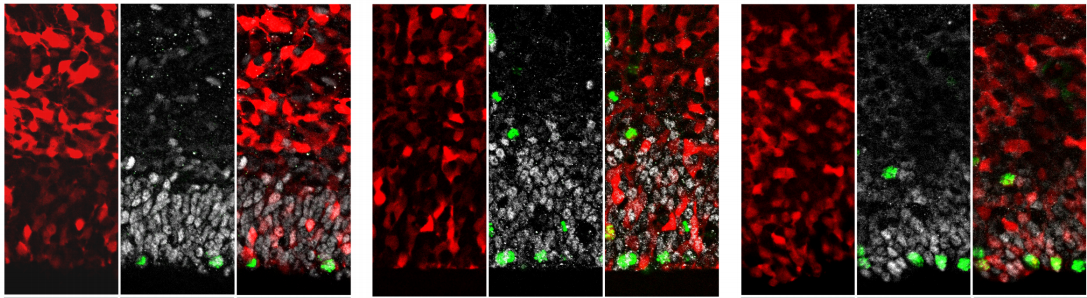
In this episode we’re going back to the very beginning, telling the stories of the midwives of the field of developmental genetics, two researchers whose work helped to reveal the secrets of life in its very earliest stages: Hilde Mangold and Salome Gluecksohn-Waelsch.
The tale of developmental genetics is a thrilling one, with everything you need for a good story. There’s politics, drama, upheaval, prejudice, and even a suspicious death. So hold on tight, this is a good one.
Full show notes, transcript, music credits and references online at GeneticsUnzipped.com.
We are inviting applications for three postdoctoral positions within the Zebrafish Neuronal Networks Group at the Department of Neuroscience (Uppsala University, Sweden). The research group, established in 2015, currently consist of four post-docs, three PhD students and a lab-manager. We investigate the formation and function of neuronal networks in zebrafish, with a focus on the locomotor network and the lateral line system. The goal is to connect the formation of a circuit with its functionality, both at the cellular and at an organism level. We use a range of techniques to reveal how cells are generated and integrated within a network and how their activity affect behaviour. For example, we employ CRISPR/Cas9 mediated knock-in to label new neuronal populations and use Light-sheet microscopy to perform in vivo time-lapse imaging to visualize their formation. To study the function of specific neurons within a circuit we use optogenetics and calcium imaging combined with analysis of animal behaviour. Furthermore, we use single cell RNA sequencing to identify subspecialization and aim to establish viral tracing to investigate the connectome of defined neuronal populations.
For more information regarding our research, see the links below:
Uppsala is a university city of 172.000 people that cater to all needs in a picturesque setting located only half an hour by train from the capital Stockholm. Uppsala University was founded in 1477 and has 46.000 students and 7.300 employees. The department offers courses in pedagogics and plenty of teaching opportunities for those aspiring an academic career. There are possibilities of entering mentoring programs and both the department and university have active post-doc associations.
The applicant
Applicants should have demonstrated outstanding performance throughout their previous studies by publications in peer-reviewed international journals. The candidate should be highly motivated, organized, and able to work independently. Strong English skills are required. Candidates should have a PhD degree and experience in one or more of the following is desired and meriting:
Zebrafish as a model system (Felasa C)
Optogenetics
Calcium imaging
Viral tracing in neurons
Single cell RNA sequencing
Data analysis using MATLAB or Python
Light-sheet, confocal, and/or 2-photon imaging
Electrophysiology
The positions are available immediately, with funding for 1 year with the possibility of an extension. Interested candidates should send their applications; cover letter including a brief statement of research interests and technical expertise, CV, publication list, as well as contact details of references directly to Dr. Boije (henrik.boije@neuro.uu.se).
The Sir William Dunn School of Pathology at the University of Oxford is looking for outstanding early career researchers seeking a stimulating and supportive environment in which to establish their research group as externally-funded fellows. We are specifically looking for researchers seeking mentoring and sponsorship to apply for career development fellowships (e.g. Wellcome Trust’s Sir Henry Dale, UKRI Future Leaders, MRC CDA, BBSRC David Phillips, CRUK Career Development Fellowship, etc). Researchers who succeed in securing a fellowship will then be invited to establish their independent group in the department, benefiting from a generous support package, comprehensive mentorship, career development training and opportunities to recruit Oxford undergraduate and postgraduate students.
Successful candidates will have an outstanding track record in any area of biomedical research, with a particular focus on the fundamental cell and molecular biology underlying disease. The Department celebrates diversity and we welcome applicants from diverse backgrounds that are currently underrepresented at the University of Oxford.
The deadline for applications is the 23rd of October. Find out more on our website.
Last October I had the great pleasure of interviewing Trudi Schüpbach for Development in Buenos Aires. We were at the Latin American Society for Developmental Biology meeting (check out the meeting report here), and over a pizza had a wide ranging discussion about her life in science, the place of genetics in developmental biology, and her plans for her retirement. The interview was published earlier this year and I’m putting it up here in case you missed it – check out Development’s interview archive for more.
Gertrud (Trudi) Schüpbach is Emeritus Professor of Molecular Biology at Princeton University. She was a Howard Hughes Medical Institute Investigator between 1994 and 2014, was elected to the National Academy of Sciences in 2005 and to the European Molecular Biology Organization as an Associate Member in 2000, and won the Society of Developmental Biology’s Edwin G. Conklin Medal in 2006. She has a long-term interest in the genetic and molecular control of oogenesis in Drosophila, work she continues in her retirement. In this interview, conducted at the Latin American Society for Developmental Biology’s 2019 meeting in Buenos Aires, she surveys her career, discusses the rich seam of science that came out of a maternal-effect screen and gives her advice for people considering a career in research.
Let’s go back to the beginning: when did you first become interested in science, and biology in particular?
I had always liked math, from a very young age. Therefore the sciences were of interest to me in general; but biology won out because it seemed the most diverse and complex. Even in high school (the ‘gymnasium’ as we called it in Switzerland), it felt that there were so many open questions that would provide lots of different possibilities for doing science. I also preferred doing experimental science, and that was a deciding factor that took me away from pure math. It was great to be able to look through a microscope and see tissues and organisms, which I had an opportunity to do at my school.
I then completed a general biology degree at the University of Zurich, and was given another great opportunity to experience actual science. For two semesters we were given lab space, and every month or two a different laboratory in the Biology department would bring in a new object for us to study. For instance, they gave us fly strains with attached-X chromosomes, and we had to figure out what was happening with their inheritance patterns. We were also given planarians and Hydra to cut into pieces. It was a lot of fun, because while we were provided with guidance, we were allowed to come up with our own questions. I think most of us who took that course ended up going into research.
So was the decision to do a PhD an obvious and easy choice?
At that time I just really loved being in the lab and doing experiments. I thought I could worry about career and money later – as long as I could afford to pursue a PhD, I would be happy. And so I ended up joining Rolf Nöthiger’s lab. At Zurich there was a tradition of working with flies and pursuing developmental genetics. For instance, Ernst Hadorn and his group had done work transplanting imaginal discs and seeing how fragments regenerated, which posed fascinating questions. Shortly after I joined the Nöthiger lab, Rolf and the rest of us students and postdocs became very interested in mosaic analysis – it was a new and exciting technique. My PhD was focused on using mosaics to analyse development of the genital disc.
The good thing about having Rolf Nöthiger as my adviser was that he was very hands off – very supportive, but he basically let us pursue our own ideas. This approach can sometimes be to the detriment of certain students, but in my case it really suited me, and it gave me the opportunity to try a lot of other things aside from my main project. When Eric Wieschaus came to the lab as a postdoc, as well as Janos Szabad, a postdoc from Hungary, we had a lot of stimulating discussions. We spent a lot of time in the lab, and on Saturdays, when the University dining room was closed, we would even cook together there. We started a new project studying mosaicism in the larval hypoderm together, which wasn’t part of my proscribed PhD. And then I started to look at ovaries – when I had to dissect the genitalia for my main project, the ovaries were always there and interesting mosaic patterns could also be observed in them, so I became really interested in how ovaries develop. Eric and Janos also had a project on germline development, which again provided for interesting discussions and comparisons. Finally, working with ovaries also spurred an interest in sex determination, which the Nöthiger lab began to be very involved in investigating, for example analysing the transformer genes. So sex determination of the germline was another project I was starting to do on the side. I usually had lots of different things going on – some, of course, were complete failures, but it taught me that it is best in science to try a number of experiments, because usually only some of them will succeed.
Given the interest in mosaicism at that time, were you influenced by the work of Antonio García-Bellido?
Antonio García-Bellido and my advisor were actually friends, as García-Bellido had been a postdoc in Ernst Hadorn’s lab, and Rolf and he had met there in Zurich. Every 2 years or so we used to have meetings – Nöthiger and García-Bellido, as well as Peter Lawrence, and each brought some students and postdocs. In this small group, everyone was into mosaics and what we could do with them, and that heavily influenced my PhD. At the time, we didn’t have molecular biology tools to follow cell determination and gene expression patterns, but using mosaicism to study cell linages and cell determination in terms of compartments was incredibly powerful. Later, other labs working with Drosophila also joined these meetings and it gave me the opportunity to meet many young scientists who became life-long friends and colleagues.
García-Bellido was a positive influence, but he was very tough. You’d go into Rolf’s office and García-Bellido was sitting there – you had to explain what you’d been doing, and he would really challenge everything you said. I think it was good for me, because Rolf was very gentle, and wouldn’t criticise us too much, not to the extent of García-Bellido. He would pepper me with questions: ‘How do you know this? How many clones have you counted? Couldn’t it be an artefact?’. He was always trying to find holes in our logic and in our approach. As well as making me extra careful about my data, it also made me think – well, I’m going to show him! I’ll come back with the data! Having grown up in Switzerland, I think it was good for me to experience a much more direct, and often somewhat aggressive, challenge.
How did you come to do your first genetic screen?
Eric had moved to his first job at Heidelberg at the European Molecular Biology Laboratory, while I stayed back in Zurich to finish my PhD. He and Christiane Nüsslein-Volhard decided to do a mutagenesis screen for embryonic lethal mutations. In order to isolate those mutations you have to set up thousands of little sub-lines. If a lethal mutation had been induced on a particular chromosome, they then took that line and studied the embryos. But there were many lines where no lethal had been induced, but nevertheless had females that were homozygous for a mutagenized chromosome, and this chromosome had the potential to carry a mutation affecting oogenesis. So I went up to Heidelberg a couple of times and dissected those flies to look for ovary phenotypes, and also looked at their eggs and embryos. It soon started to look really interesting, as there were a few mutations with specific phenotypes.
When the Heidelberg time neared its end, it was clear to Eric and me that we would like to be together somewhere, and he wanted to move back to the USA. We decided it would be interesting to carry out large scale maternal-effect screens: Christiane’s lab would focus on the third chromosome, and I would focus on the second chromosome, and then we would exchange mutants.
In Princeton, after using a postdoc stipend I’d got from Switzerland to do the screen, I then needed to apply for grants. I was very lucky to get a National Science Foundation grant in 1985. I was not yet a faculty member but in a ‘Research Biologist’ role, and while I had isolated a number of mutants that looked interesting, nowadays that of course would not be enough – you need more to prove that the project is really going to work and provide fundamental new insights. So it was really nice that NSF took that risk with me and gave me that grant to establish my own group. Two years later, after Jim Price, my first postdoc, had shown that the gene that we called torpedo encoded the Drosophila homolog of the EGF receptor, we could apply for National Institutes of Health grants, and the lab could grow further.
Your lab’s work was defined by the mutations that came out of the screen, particularly with regard to the dorsoventral patterning of the egg. But I guess you had no idea where they were going to lead you?
That is true, the genes corresponding to the mutants could have encoded any type of protein. We had some luck with timing: at the time of the screens, molecular biology began to become possible in whole organisms. At the start of my PhD, to do molecular biology in higher organisms was only possible for genes that produced very highly expressed proteins like histones or haemoglobins. I remember one day going with my advisor to a person at the university who was establishing a molecular biology laboratory. We told him we wanted to find the molecular nature of some of the genes we were working on – how could we do this? He basically told us it was going to be impossible to clone developmental genes in Drosophila. But then, just 10 years later, people had invented methods like chromosomal walking, and if you had enough chromosomal aberrations such as breakpoints within your gene you could look at the salivary glands and pin down the location of the gene within your molecular walk. It really was transformative, and soon the methods became even more powerful combined with the wealth of other genetic resources we already had in Drosophila.
When you do a mutagenesis screen and end up with so many lines, you have to decide which ones to work on. I think that in the case of gurken (which we later found encodes a TGFα-like protein), the ventralisation phenotype was so striking, reliable and intriguing, and the mutants were so fully penetrant, that you could both believe in the gene’s importance and work easily with it. Because of my prior exposure to mosaic analysis, I tested a lot of the genes to see whether they were required in the germline or the follicle cells. gurken was required in the germline but torpedo (which we and Benny Shilo’s lab later found encodes the EGF receptor) was required in the follicle cells. So that finding stood out: you could get ventralised eggs when either the germline (gurken) or the follicle cells (torpedo) were affected. It pointed to a communication between those two cell types that was necessary to establish the whole dorsoventral pathway for the organism.
In the early 1990s I had two postdocs – Shira Neuman-Silberberg, who was cloning gurken, and Siegfried Roth who was cloning cornichon, a gene he’d identified while in Christiane Nusslein-Volhard’s lab (gurken means cucumber in German, and cornichon does in French). When gurken turned out to encode TGFα it was immediately clear how it functioned, whereas cornichon encoded a protein that had something to do with secretion of Gurken, and was in a way less special, a ubiquitously expressed ‘helper’ protein. But Siegfried is an excellent scientist and he had noticed that that the phenotypes of gurken and cornichon included a duplication of the anterior follicle cell fates at the posterior of the egg, leading to a second micropile. He decided to investigate this further and pretty quickly found out that the true null mutations in both genes also caused aberrations in the anterior-posterior axis formation, which led to an important insight on axis formation during oogenesis, and he ended up with a very nice publication. It is a good example for how an initially unpromising story can turn around and blossom into something important, if you’re paying attention to what may initially look like an unexplained curiosity.
We also had other ventralising mutants which had a more variable phenotype, and it turned out they were DNA repair enzymes. This made them initially less obviously interesting, and frankly, baffling, but fortunately a graduate student, Amin Ghabrial, was not daunted by this finding – he insisted on trying to figure out why DNA repair enzymes would have a ventralising phenotype. He completed a really nice story showing that the problem with the mutants arose in meiosis, when the oocyte needs these repair enzymes. If damaged, broken DNA is not repaired efficiently, there is a block on development activated by a damage checkpoint, and because of that block, Gurken protein is not produced at the right time and to the right amount. Eventually this block is somehow relieved, egg development resumes, but now it’s too late for correct dorso-ventral patterning of the egg. Hence, one sees these variable, ventralised phenotypes. Without the mutants, we would never have thought of meiotic DNA repair as being a prerequisite in egg patterning.
How important has modelling been in your investigation of egg patterning?
It’s been very informative. Some years ago I met a young colleague, Stas Shvartsman, who came to Princeton working in the Department of Chemical Engineering. His background was in modelling of signalling pathways, like the ERK pathway, and he came to our lab interested in whether he could model something in egg patterning. He didn’t know much about flies or genetics, and I didn’t know anything about modelling or coding, but we soon found some brave graduate students who went back and forth between the labs and learned to do both. It was very enriching to our research. People coming from such a different field ask questions that initially you think are very naïve – ‘how many molecules of this factor are released?’ or ‘what’s the concentration of that particular protein?’. You think, well, of course we can’t measure that in flies. But why not? And would there be a way to estimate, or at least approximate it? And often you realise genetics can help here because it allows you to do things like halving the dose of a receptor by using a chromosomal deficiency: then you can measure how this affects the signalling response. Your model then has to be able to correctly predict the outcome. Such quantitative experiments can tell you a lot about directness, feedback, compensation mechanisms and so on. All of these processes have become really interesting to many of us working in development. It is always fun in science to be confronted with something new that you initially don’t know anything about – you can feel like a graduate student again, which I think is a great privilege.
“It is always fun in science to be confronted with something new that you initially don’t know anything about”
Given how helpful modelling has been for you, can genetics alone only get you so far in understanding development?
Yes, I agree, there is a limitation. This then raises the question of the position of genetics in the field. Nowadays we do so much genomics, and you can get long lists of transcripts in different tissues, and now even from single cells. But once you get this list, how do you know which ones are the important ones? Even if you can refine your candidates, it is usually still a long list, and then mutant analysis is vital. So there will always be a need for genetics to test things out, even though we are now doing genetics differently – CRISPR, morpholinos, RNAi. It’s still a very strict test of functionality.
More broadly, which big questions will dominate developmental biology in the next decade or two?
We’re at a very exciting point at the moment, with all these new techniques at our disposal. Microscopy has come such a long way – it allows us to ask new questions, and there’s still a very rich field that we can explore. One question is mechanics in development: we know a little in terms of actin and myosin for instance, but there’s so much left to find out. And then combine that with the possibility of single cell transcriptomics, where we’re really at the beginning. People will say we found 16 cell types in a tissue during development, but are they really 16 distinct cell types? Or are the cells going through phases and stages, and have you just taken a snapshot? Is all gene expression meaningful? These are very exciting questions to ask. Even in the Drosophila embryo, you might think everything is known, all of these genes have been cloned and their expression patterns analysed, and so on. But you can now visualise transcription live in the embryo, and ask about the dynamics of a process that was previously just seen in snapshots.
I understand you’re recently retired: are you still staying in touch with science?
I’m now officially retired as a faculty member in the university. I still have a lab space and an office, but no longer have graduate students or postdocs, even though I’m still involved in collaborations with other people’s students. I’m finding it quite fun actually not to have that much responsibility. As a PI you hope that every one of your graduate students and postdocs has a wonderful experience and gets great papers and goes on to find a wonderful job. And sometimes this just doesn’t quite work out, so you ask yourself was it my fault, did I give them the wrong project? It’s not like anyone in my lab really failed, but it is a bit of a relief not to have to worry about that responsibility.
I’m still doing research – in particular I have a number of mutants that haven’t been well characterised and which I’d like to look at a little bit more. I’ll investigate whether they are going to be interesting and reliable, and maybe they could be a project that could be handed off to someone later. You can’t just give someone a mutant, but they might take it on if you can make the case for why it would be interesting to pursue. And then these orphan mutants would find a home!
Do you have any advice for someone considering a career in developmental biology?
I always tell undergraduates to read widely and find something that interests them. The best thing to do is follow some problem that fascinates you, and only think about your career and so on after the PhD. I also think as a PhD student you should aim to start out with the best conditions possible to figure out whether you like research or not – do you have fun, are you happy in the lab? Finding a good lab and an adviser who fits your personality is essential, and don’t worry too much about whether it is this organism or that, or whether it’s related to cancer or some eminently fundable problem.
After the PhD, people tend to have a good sense of what they like to do. For instance, I’ve had really wonderful graduate students who come to the end and tell me they don’t want to work at the bench any more, they just don’t enjoy that type of work any longer and want to do something else. By that time they know themselves better – the PhD hopefully has given the students the opportunity to really explore what excites them and matches their talents. It also comes back to choosing the right lab: if you’re in a miserable place and you don’t like your advisor, well of course you won’t want to carry on. But if the experience has been good, the student knows what they like best about science, and what types of problems fascinate them, then it’s a question of channelling all of this into a new project and a new laboratory and making it work.
Is there anything Development readers would be surprised to find out about you?
Since I have three daughters, there was not that much time between lab, teaching and parenting to take up many hobbies. Actually that’s one of the things I’m looking forward to now that I have fewer responsibilities. For example, I’d love to play more of the piano, and would like to travel more and not only for work. I like languages and I want to pursue that a little bit more – Spanish would be a great place to start, as I’m appreciating here in Buenos Aires.
But on the topic of family, I often find that young women scientists are worried about whether they will be able to combine family and work. And so I’d just like to say: yes, you can combine these things successfully. Our three daughters are happy young people and don’t seem to have held any grudges against us, even though we were not always both there for their school events or sports games. Having a supporting partner, of course, helps immensely. In the end, our daughters say that they are actually glad we were not hovering over them every minute, they feel it allowed them to become more independent. So while every family has, of course, to find their own balance, it should not be a hindrance for either parent to be pursuing science at a high level.
Kamberov lab at the University of Pennsylvania Perelman School of Medicine, Philadelphia PA.
About the lab: The primary research focus of the Kamberov lab is to define the developmental programs controlling the formation and patterning of skin organs like sweat glands and hair follicles and to discover how these programs have been modified during to generate diversity in skin traits among modern humans as well as to give rise to human-specific skin traits . To this end, the lab leverages powerful genetic and genome editing tools for discovery and functional testing in the mouse coupled with in vitro study in mouse and human cell culture, comparative evolutionary genomics and high-throughput single transcriptomics.
About the position: A post-doctoral position is available leading a project on mechanisms underlying developmental specificity and evolutionary variation of enhancers controlling the formation and patterning of skin appendages. The position is supported by NIH funding.
To apply: Interested applicants should send a CV, contact information of three references and a letter detailing your interest in the position to Yana Kamberov. Applicants should hold a Ph.D. in biology or related field. Applicants with a background in developmental biology, enhancer biology, mouse genetics, regenerative biology or evolutionary genetics are especially encouraged to apply.
There are two open positions in my lab. We are the Neural circuits and Evolution lab at The Francis Crick Institute in London (https://prietogodinolab.org/). The lab employs a multidisciplinary approach to understand how neural circuits function and evolve, combining techniques that range from in vivo calcium imaging, electrophysiology, electron microscopy circuit tracing, molecular biology, single-cell RNA sequencing, genome editing, behavioural analysis and field work. The two open positions are:
Senior Lab Research Scientist. This position is best suited for a motivated postdoctoral researcher that wishes to continue doing amazing science in a collaborative environment with a permanent contract, rather than following the PI route. The role will be a mix of a permanent advanced postdoctoral researcher and lab manager supporting other people’s projects in the lab. The link to the official application and job description can be found here: https://my.corehr.com/pls/frckrecruit/erq_jobspec_version_4.display_form?p_company=1&p_internal_external=E&p_display_in_irish=N&p_applicant_no=&p_recruitment_id=014727&p_process_type=&p_form_profile_detail=&p_display_apply_ind=Y&p_refresh_search=Y
Postdoctoral researcher. This position is best suited for a motivated finishing PhD student or a postdoc that wishes to continue their academic path in a supportive environment, it is a four year contract with possibility of extension to a total of 6 years. In addition to a supportive lab environment, the Crick provides an excellent postdoctoral training programme, and we have regular meetings as well as formal and informal collaborations with other neuroscience groups at the crick. The link to the official application can be found here:https://my.corehr.com/pls/frckrecruit/erq_jobspec_version_4.display_form?p_company=1&p_internal_external=E&p_display_in_irish=N&p_applicant_no=&p_recruitment_id=014747&p_process_type=&p_form_profile_detail=&p_display_apply_ind=Y&p_refresh_search=Y
Both positions require independence and a strong interest in either cellular and molecular neuroscience or circuit/systems neuroscience and evolution.
Informal enquires can be made to lucia.prietogodino@crick.ac.uk
I would be thankful if you could distribute this adds widely.
PhD offers: modelling the effect of noise in gene regulatory network governing early mammalian development, starting from 1 October 2020 or later – (1 +3 years )
We are seeking two PhD students to work in the Unit of Theoretical Chronobiology (Brussels, Belgium) on a project related to the modelling of cell differentiation. This interdisciplinary project will focus on the relation between the structure of the gene regulatory network (GRN) governing cell differentiation and its sensitivity to noise.
For more information, see: https://www2.ulb.ac.be/sciences/utc/home.html
The Tissue Regeneration Laboratory within the Biology Department at Wake Forest University is seeking a qualified and highly motivated individual for the position of Lab Manager / Research Assistant to support our work uncovering mechanisms of tissue regeneration in axolotl salamanders. The primary tasks will include managing lab resources, caring for the lab’s axolotl colony, and undertaking research. The candidate will be involved in setting up our lab as it launches at Wake Downtown. Wake Downtown is located in Winston-Salem’s Innovation Quarter, adjacent to the Wake Forest School of Medicine and the Wake Forest Institute for Regenerative Medicine (WFIRM). Winston-Salem boasts a phenomenal cost of living, a thriving arts scene as the home to the North Carolina School of the Arts, and is only a few hours’ drive to either the Outer Banks or the Blue Ridge Parkway/Smoky Mountains.
The candidate will be responsible for managing lab inventories, implementing safety protocols, maintaining and caring for axolotls and their facility, and conducting lab work such as PCR genotyping, RNA and DNA extractions, molecular cloning, creating transgenic axolotls, and fluorescence microscopy. The candidate should be motivated to gain proficiency at troubleshooting experiments, analyzing data, and communicating results. Opportunities exist for independent research and mentorship toward next professional steps (e.g. postbac to grad/med school). The candidate should minimally have a Bachelor’s degree and a fundamental understanding of molecular biology, including hands-on research experience in cell and molecular biology or a related field. Experience working with animals, especially aquatic organisms, is appreciated but not required. Excellent organizational, record-keeping and collaborative interpersonal skills are essential.
This position has full benefits and is funded for two years, with the possibility of extension dependent upon lab funding. Salary will be commensurate with prior experience. We are intentional about building a diverse, inclusive, and supportive team of colleagues. More information about our lab values can be found here.
To apply, please send a CV, a cover letter expressing interest and highlighting relevant experience, and contact information for 3 references to Dr. Josh Currie (currie.regenerationlab@gmail.com). Please reference “Tissue Regeneration Lab Manager / Research Assistant” in the subject line. Starting dates are flexible beginning from September 1st, 2020.
The N-Glycome regulates the endothelial-to-hematopoietic transition
Dionna M. Kasper, Jared Hintzen, Yinyu Wu, Joey J. Ghersi, Hanna K. Mandl, Kevin E. Salinas, William Armero, Zhiheng He, Ying Sheng, Yixuan Xie, Daniel W. Heindel, Eon Joo Park, William C. Sessa, Lara K. Mahal, Carlito Lebrilla, Karen K. Hirschi, Stefania Nicoli
Maternal iron deficiency perturbs embryonic cardiovascular development
Jacinta I. Kalisch-Smith, Nikita Ved, Dorota Szumska, Jacob Munro, Michael Troup, Shelley E. Harris, Aimée Jacquemot, Jack J. Miller, Eleanor M. Stuart, Magda Wolna, Emily Hardman, Fabrice Prin, Eva Lana-Elola, Rifdat Aoidi, Elizabeth M. C. Fisher, Victor L. J. Tybulewicz, Timothy J. Mohun, Samira Lakhal-Littleton, Eleni Giannoulatou, Duncan B. Sparrow
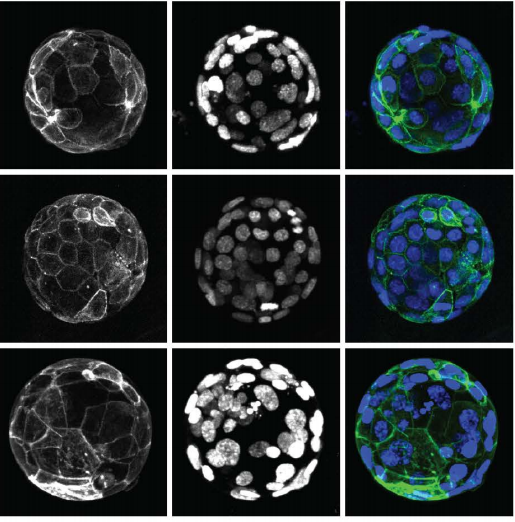
Cell surface fluctuations regulate early embryonic lineage sorting
Ayaka Yanagida, Christopher Revell, Giuliano G. Stirparo, Elena Corujo-Simon, Irene M. Aspalter, Ruby Peters, Henry De Belly, Davide A. D. Cassani, Sarra Achouri, Raphael Blumenfeld, Kristian Franze, Ewa K. Paluch, Jennifer Nichols, Kevin J. Chalut
Epithelial layer unjamming shifts energy metabolism toward glycolysis
Stephen J. DeCamp, Victor M.K. Tsuda, Jacopo Ferruzzi, Stephan A. Koehler, John T. Giblin, Darren Roblyer, Muhammad H. Zaman, Scott T. Weiss, Margherita DeMarzio, Chan Young Park, Nicolas Chiu Ogassavara, Jennifer Mitchel, James P. Butler, Jeffrey J. Fredberg
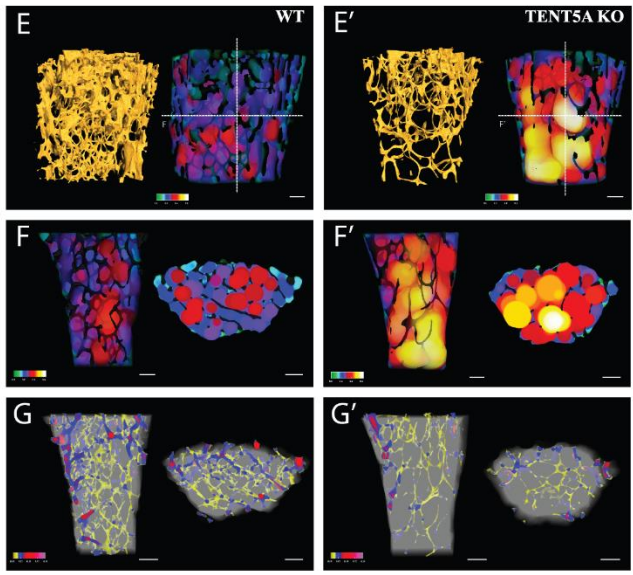
Cytoplasmic polyadenylation by TENT5A is required for proper bone formation
Olga Gewartowska, Goretti Aranaz Novaliches, Paweł S Krawczyk, Seweryn Mroczek, Monika Kusio-Kobiałka, Bartosz Tarkowski, Frantisek Spoutil, Oldrich Benada, Olga Kofroňová, Piotr Szwedziak, Dominik Cysewski, Jakub Gruchota, Marcin Szpila, Aleksander Chlebowski, Radislav Sedlacek, Jan Prochazka, Andrzej Dziembowski
Paranode stability requires UNC5B expression by oligodendrocytes
Omar de Faria Jr., Diane S. Nakamura, Samuel Clemot, Doyeun Kim, Mihai Victor Mocanu, Roland Pilgram, Jenea M. Bin, Edwin W. Wong, Amir Shmuel, Abbas Sadikot, Susan L. Ackerman, Timothy E. Kennedy
Cell-type, single-cell, and spatial signatures of brain-region specific splicing in postnatal development
Anoushka Joglekar, Andrey Prjibelski, Ahmed Mahfouz, Paul Collier, Susan Lin, Anna Katharina Schlusche, Jordan Marrocco, Stephen R. Williams, Bettina Haase, Ashley Hayes, Jennifer G. Chew, Neil I Weisenfeld, Man Ying Wong, Alexander N. Stein, Simon Hardwick, Toby Hunt, Zachary Bent, Olivier Fedrigo, Steven A. Sloan, Davide Risso, Erich D. Jarvis, Paul Flicek, Wenjie Luo, Geoffrey S. Pitt, Adam Frankish, August B. Smit, M. Elizabeth Ross, Hagen U. Tilgner
Wnt/PCP-primed intestinal stem cells directly differentiate into enteroendocrine or Paneth cells
Anika Böttcher, Maren Büttner, Sophie Tritschler, Michael Sterr, Alexandra Aliluev, Lena Oppenländer, Ingo Burtscher, Steffen Sass, Martin Irmler, Johannes Beckers, Christoph Ziegenhain, Wolfgang Enard, Andrea C. Schamberger, Fien M. Verhamme, Oliver Eickelberg, Fabian J. Theis, Heiko Lickert
Human muscle stem cells are refractory to aging
James S. Novak, Davi A.G. Mázala, Marie Nearing, Nayab F. Habib, Tessa Dickson, Olga B. Ioffe, Brent T. Harris, Marie N. Fidelia-Lambert, Christopher T. Rossi, D. Ashely Hill, Kathryn R. Wagner, Eric P. Hoffman, Terence A. Partridge
Macrophages provide a transient muscle stem cell niche via NAMPT secretion
Dhanushika Ratnayake, Phong D. Nguyen, Fernando J. Rossello, Verena C. Wimmer, Abdulsalam I. Isiaku, Laura A. Galvis, Alasdair J. Wood, Ziad Julier, Thomas Boudier, Viola Oorschot, Kelly L. Rogers, Mikaël M. Martino, Christophe Marcelle, Graham J. Lieschke, Jeroen Bakkers, Peter D. Currie
Hematopoietic stem cells fail to regenerate following inflammatory challenge
Ruzhica Bogeska, Paul Kaschutnig, Malak Fawaz, Ana-Matea Mikecin, Marleen Büchler-Schäff, Stella Paffenholz, Noboru Asada, Felix Frauhammer, Florian Buettner, Melanie Ball, Julia Knoch, Sina Stäble, Dagmar Walter, Amelie Petri, Martha J. Carreño-Gonzalez, Vinona Wagner, Benedikt Brors, Simon Haas, Daniel B. Lipka, Marieke A.G. Essers, Tim Holland-Letz, Jan-Philipp Mallm, Karsten Rippe, Paul S. Frenette, Michael A. Rieger, Michael D. Milsom
A Wnt-specific astacin proteinase controls head formation in Hydra
Berenice Ziegler, Irene Yiallouros, Benjamin Trageser, Sumit Kumar, Moritz Mercker, Svenja Kling, Maike Fath, Uwe Warnken, Martina Schnölzer, Thomas W. Holstein, Markus Hartl, Anna Marciniak-Czochra, Jörg Stetefeld, Walter Stöcker, Suat Özbek
The Echeverri lab at the MBL seeks a highly motivated individual to join the Eugene Bell Center for Regenerative Biology and Tissue Engineering as a Postdoctoral Researcher. The successful candidate will work on the molecular mechanisms of scar free skin regeneration in axolotls.
The specific goal of the project is to examine the role of different cell types in responding to the injury cues and in later remodeling collagen. The position will initially be offered for two years with the possibility of extension.
Basic Qualifications:
Applicants should have a Ph.D. in a biology related field. Must have prior experience working in the field of cell and developmental biology, as well as experience with molecular biology, cell culture and research animals. Must be independent, enthusiastic, self-motivated, productive, and enjoy working in a collaborative environment.
Preferred Qualifications:
The ideal candidate will have direct experience with working in vivo in an animal model. Previous experience with cell culture, molecular biology and imaging would be a plus.
Required documents:
1. Cover letter explaining specifically why you are interested in joining our lab to work on this project and what positive qualities you would bring to our team.
2. Curriculum vitae.
3. Apply online : https://recruiting.ultipro.com/MAR1033MBL/JobBoard/4c3007c3-6354-41de-a13f-d95be60d91e9/?q=&o=postedDateDesc
4.
 In this episode we’re going back to the very beginning, telling the stories of the midwives of the field of developmental genetics, two researchers whose work helped to reveal the secrets of life in its very earliest stages: Hilde Mangold and Salome Gluecksohn-Waelsch.
In this episode we’re going back to the very beginning, telling the stories of the midwives of the field of developmental genetics, two researchers whose work helped to reveal the secrets of life in its very earliest stages: Hilde Mangold and Salome Gluecksohn-Waelsch. (No Ratings Yet)
(No Ratings Yet)