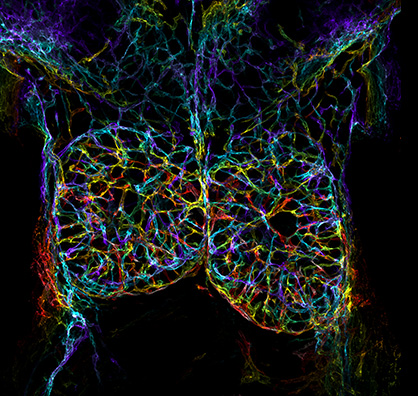
Are you keen to get more science communication experience? Is your research related to developmental and stem cell biology? The Node is looking for a reporter to attend and report from the Biologists @ 100 conference, happening 24-27 March 2025 in Liverpool, UK. This conference is a celebration of the 100th birthday of The Company of Biologists, bringing together different communities, including cell, developmental, experimental and disease biologists, and will incorporate the Spring Meetings of the British Society for Cell Biology (BSCB) and the British Society for Developmental Biology (BSDB). Registration for the conference is open and the abstract deadline is 13 December.
The selected reporter will have their conference registration fee waived (but will have to cover their own transport and accommodation costs*). They will be expected to attend the full conference. While the reporter will mainly focus on reporting from the ‘Cell and developmental biology’ scientific track, they are welcome to attend sessions from other tracks in the programme.
If you are interested, we’d love to read about your ideas of what you’d do as a reporter at the conference. If selected, we will work with you to develop your ideas and assist you at the conference.
If you have any questions about the role, don’t hesitate to email us at thenode@biologists.com.
A proposal (max 200 words) with idea(s) of what you would do as a conference reporter. We are open to suggestions of different formats of reporting. Please include a rough output you expect to produce from the reporting, and explain how the content will appeal to the Node readers.
A paragraph (max 200 words) telling us about yourself, your research, and any relevant science communication experiences you have.
Application deadline: Friday 29 November 2024
Our sister community site FocalPlane is also looking for a conference reporter. If you want to focus on reporting on cell biology and the use of microscopy in research, head over to their website to apply.
*Note: we encourage you to apply for conference grants to cover the transport/ accommodation costs, such as grants from BSDB and DMM.
The Indian Society of Developmental Biologists is the national society that represents developmental and stem cell biologists across India. We are pleased to introduce the society and our activities to the Node readers. Reading on, you will find some of our recent initiatives and posts. Do visit the InSDB website to check out more. You can get in touch with us at info@insdb.in if you would like to contribute/be a volunteer for InSDB.
Gulmohar-Postdoc Talk Series
Gulmohar is our new, virtual, post-doc talk series through which we aim to bring together early-career scientists from around the world to share their research. This is a chance to hear from the next crop of PIs about their work, where their research is heading, and what’s next for them. Whether you are starting your research career, or already knee-deep in it, through this series, you will get to know about the different fields within the broad scope of developmental biology research. After every session, we also plan to have a post-talk informal discussion where you get to connect with the speakers and receive insight into their scientific journey and potential opportunities for your own.
We are currently accepting applications from postdocs who would like to be a speaker for the series. If you fit the bill, please sign up here, and we will get in touch with you.
Science Simplified
Science Simplified is our series where we pick up a recently published research paper in developmental biology and distill it to the liking of a general audience. You can read the first article in this series here.
InSDB is also accepting articles from our audience! These could be stories on the latest research from the stem cell and developmental biology fields and perspectives on matters that are relevant to our audience. Anyone is welcome to pitch stories to us. We accept articles that are thoroughly researched and convey the excitement, relevance, and fascination of the subject to any curious reader. If you are interested in science writing and would like to sharpen your skills, this is your cue to get your pen and paper out! Also, do read our submission guidelines to know more. If you would like to contribute, please add your entries here.
Behind The Bench
Behind the Bench is our interview series where we feature InSDB members, their stories, and the science they do. In each episode, we sit down with a member to know about their journey in science, from their early inspirations to the serendipitous moments along the way. Through this series of interviews, we aim to highlight the members who make our society what it is today. You can read these interviews here.
—
If you would like to follow our updates closely, we are on X, LinkedIn, and Instagram!
Daniel Riveline,1 Karsten Kruse,2 Mukund Thattai,3 Guntram Bauer4
Scientific communication was pioneered in the 17th century (Ref. 1). Scientists were called ‘sçavans’and had to embrace the humanities and the sciences to present their discoveries with clarity. These achievements are difficult nowadays, with the detailed technical knowledge involved in biology, physics, chemistry and mathematics research. In this context, it is challenging to present the depth of each side of an interdisciplinary scientific question, and often one field is featured more prominently, to the detriment of the other. To address this issue, we organised a Symposium in which presentations were given as tandem talks, with participants from biology and physics or from chemistry and biology. Topics ranged from cell positioning in the embryo (Eric Wieschaus and William Bialek), to liquid condensates (Anthony Hyman and Frank Jülicher), to the physics of photosynthesis (Donatas Zigmantas and Thomas Renger). We found this tandem format to be extremely successful (see Symposium online Ref. 2). Concepts from physics, biology and chemistry were explained in accessible yet informative language, and the audience could see how interdisciplinary engagements developed. Importantly, the speakers highlighted active points of debate and articulated unresolved issues, leaving space for new breakthroughs and scientific directions. As a metaphor for collaboration, a musical duo on the violin and cello illustrated how each voice in a partnership could be heard. Using a Bach counterpoint as the paradigm, the musicians from the Philharmonic Orchestra of Strasbourg explained how their pieces were harmonious only with shared and alternating voices.
References:
1- A History of Scientific Journals, Aileen Fyfe, Noah Moxham, Julie McDougall-Waters, Camilla Mørk Røstvik, UCL Press London (2022) https://www.uclpress.co.uk/products/187262
1- Laboratory of Cell Physics, IGBMC, CNRS and University of Strasbourg, Strasbourg, France, riveline@unistra.fr
2- Departments of Biochemistry and Theoretical Physics, University of Geneva, 1211 Geneva, Switzerland, Karsten.Kruse@unige.ch
3- Simons Centre for the Study of Living Machines, National Centre for Biological Sciences, Tata Institute of Fundamental Research, Bengaluru, India, thattai@ncbs.res.in
4- The International Human Frontier Science Program Organization, 12 Quai Saint-Jean, 67080 Strasbourg, France, gbauer@hfsp.org
Oxford Open Doors is an annual event that offers the public a unique opportunity to explore the historic and modern buildings of Oxford, many of which are normally closed to the public. Organized by Oxford Preservation Trust, the event takes place over a weekend in September and allows visitors to experience the architectural and cultural heritage of the city first hand.
As part of this year’s Oxford Open Doors event, researchers from the Institute for Developmental and Regenerative Medicine (IDRM) invited the public into their labs, offering a rare glimpse into their cutting-edge research. Visitors were guided through research areas and imaging facilities by enthusiastic staff eager to share their work.
The IDRM is a unique flagship institution at the University of Oxford, dedicated to meeting an ambitious challenge: two thirds of all deaths world-wide are due to non-communicable diseases, many of which are cardiovascular, neurological or immune system disorders that have a developmental origin, representing an urgent unmet clinical need. The mission of the IDRM is the development of new drugs and therapeutic strategies to tackle these chronic illnesses.
At its core is a formal merger of developmental biology and regenerative medicine in the form of over 200 cardiovascular, neuroscience and immunology scientists, integrating their expertise to foster multidisciplinary collaborations.
Under this research interest, we’ve developed a number of initiatives for Public Engagement that we deployed at this Open Day: the first, Synchrony, is a unique science-art project created in collaboration with computational artist Andy Lomas. Part of the public engagement efforts of the Wellcome funded Human Developmental Biology Initiative, Synchrony brings the complex world of developmental biology to life.
Researcher Matthew Stower talking to visitors about the HDBI research and Synchrony stereo viewer. copyright 2024 Tomoko WatanabeSynchrony stereo viewer. copyright: Andy Lomas
Synchrony allowed attendees to interact with a 3D artificial simulation of growth and pattern formation, changing the rules that govern the formation of some of the fascinating shapes that we can observe during embryonic development. This immersive experience captured visitors’ curiosity and sparked conversations about the intricate biological processes that direct embryo development and organ formation with Matthew Stower who represented the HDBI researchers. The creative process itself involved lengthy discussions between Andy and researchers Matthew Stower, Shifaan Thowfeequ, Claudio Cortes, Tomoko Watanabe and Shankar Srinivas.
Mouse embryonic thymus. copyright David Grainger
In addition to Synchrony, visitors also engaged with scientific discovery through SyGlass, a virtual reality software for exploring, annotating and sharing 3D data. SyGlass allows users to fully immerse themselves in stunning microscopy images, rendered in interactive 3D in a VR environment. A breath-taking image of the embryonic thymus, contributed by David Grainger, enabled attendees to explore the beauty and intricacies of the 3D structure of this organ. Both children and adults delighted in slicing through the tissue sections and navigating the complexities of the thymus in real time.
copyright Tomoko Watanabe
We would like to thank The Node for their support. Your extremely popular postcards gave us the perfect ‘hook’, to talk about the wonders of development and how it takes place across species. These visuals helped visitors connect the beauty and complexity of developmental biology with the cutting-edge research happening within IDRM, while at the same time sparking very interesting conversations about research and ethics.
Núria Ros-Rocher1, Júlia Ramon Mateu2, Elizabeth Williams3
1 Institut Pasteur, Université Paris-Cité, CNRS UMR3691, Evolutionary Cell Biology and Evolution of Morphogenesis Unit, 25-28 Rue du Dr. Roux, 75015 Paris (France)
2 Laboratoire de Biologie du Développement de Villefranche-sur-mer (LBDV), UMR7009 CNRS/UPMC, Institut de la mer de Villefranche, Quai de la Darse, 06234 Villefranche-sur-Mer Cedex, France
3 University of Exeter Biosciences, Streatham Campus, Stocker Rd, Exeter EX4 4QD, UK
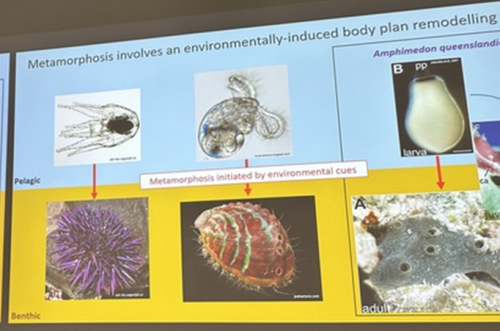
With some beautifully warm and sunny mid-summer weather, attendees of the EuroEvoDevo2024 gathered on Friday morning 28th June at the University of Helsinki for a symposium on ‘Environmental cues for life cycle transitions’. This was one of twenty-four symposia at this year’s meeting, with a record attendance of over 600 people. It was co-organized by Vincent Laudet (Okinawa Institute of Science and Technology, Japan and Academia Sinica, Taiwan) and Elizabeth Williams (University of Exeter, UK) and involved ten presenters. This symposium aimed to examine life cycle transitions and the influence of the environment on them from a broad perspective across organisms from different kingdoms of life. What aspects of the environment trigger changes in an organism’s form and physiology and how do organisms sense these?
Although starting early on the final morning of the conference, our enthusiastic late-night discussions at the nearby conference pub, the ‘Thirsty Scholar’, were offset by strong Finnish coffee, a selection of fresh vegetarian foods, the promise of the conference dinner and party later that evening, and our shared interest in the evolution of complex life cycles. The symposium brought together diverse researchers and topics, with study organisms ranging from liverwort to diatoms, choanoflagellates, insects, marine invertebrates, and fish. Environmental cues discussed ranged from abiotic factors such as light, temperature and salinity, to biotic cues including microbial biofilms, coralline algae, and food type and availability.
The symposium’s introductory slide included example images of the diverse organisms studied by presenters.
Overall, five different types of life cycle transition were discussed. Arild Husby, from Uppsala University (Sweden), presented recent work on morphological polyphenism in water striders, which is induced by changes in photoperiod. When exposed to more hours of light in the day, water striders will develop short-wing phenotype and under short day lengths will develop long-winged phenotypes. RNA sequencing followed by RNAi knockdown and ELISA measurements of the insect moulting hormone ecdysone indicated that this polyphenism is regulated by the Fat/Hippo pathway (Gudmunds et al. 2024), with a possible role for ecdysone signalling. The Fat/Hippo pathway controls organ size in animals and relates to a fundamental question in developmental biology: how does an organ know when to stop growing after reaching a particular size? The ecdysone signalling pathway was also the focus of work presented by Giulia Campli, from the University of Lausanne, Switzerland, who investigated the moulting gene repertoire across an impressive 153 proteome dataset, comprehensive of insect, crustacean and chelicerate arthropod species, highlighting changes and novelties in the molecular machinery driving the moulting process, a second type of life cycle transition for exoskeleton renewal (Campli et al. 2024).
A third common type of life cycle transition among animals is metamorphosis from a larval to a juvenile form. Several speakers focused on this transition across a wide range of animals, from sponges to fish. Océane Blard, PhD student at the University of Queensland (Australia), presented remarkable brightfield timelapses and fluorescent confocal images illustrating the morphological and cellular changes observed during the first 6 hours of metamorphosis in the marine sponge Amphimedon queenslandica. She also tracked the fate of larval sensory-secretory cells, known as flask cells, which are proposed to detect cues from coralline algae to initiate settlement and metamorphosis (Nakanishi et al. 2015). These cells appear to be able to transdifferentiate, and future research will focus on understanding the molecular mechanisms underlying this process using single cell transcriptomics. Elizabeth Williams, research fellow at the University of Exeter (UK), focused on understanding how free-living larvae settle and metamorphose to become seafloor-dwelling juveniles. Elizabeth and colleagues found that microalgae biofilms, notably diatoms, induce a settlement response in the marine worm Platynereis dumerilii (Hird et al. 2024), they described similar sensory-neurosecretory cells across polychaete, cnidarian and mollusc larvae, potentially responsible for timing metamorphosis according to external environmental conditions. In future work, Elizabeth and colleagues will focus on the morphology, function and molecular signalling mechanisms of these cells.
Océane Blard identified conserved mechanisms of metamorphosis between sponges and other animals
Júlia Ramon Mateu, PhD student at the Laboratoire de Biologie du Développement (LBDV) in Villefranche-sur-mer, presented her PhD work on the cellular and molecular mechanisms of cnidarian settlement. She identified and characterized the cell types forming the aboral end (head) of the planula larva of three cnidarian species belonging to the two main clades of Cnidaria: the jellyfish Clytia hemisphaerica and the corals Astroides calycularis and Pocillopora acuta. To do so, she generated bulk RNA-seq data from the planula aboral end and single-cell RNA-seq data from the same larval stage for each species. Through a comparative analysis of transcriptomic profiles, morphology and distribution of aboral cell types between the larvae of Clytia and the two coral species, she defined the common cellular architecture of the planula aboral end featuring two broad cell-type classes: secretory-like and neural-like. Of particular interest, she identified genes related to taurine uptake and catabolism as a shared feature of specialized cells in the planula aboral end: associated with a neural cell type in Clytia and a specialized epidermal cell type in the two coral species. Using settlement assays developed for both Clytia and Astroides planulae, she could show that exogenous taurine delays or prevents settlement in both species. Júlia concluded her presentation with an updated model of the cells and molecules involved in cnidarian larval settlement, proposing taurine uptake as a key factor in the regulation of the settlement response.
Vincent Laudet (Okinawa Institute of Science & Technology, Japan and Academia Sinica, Taiwan) also investigates the environmental influences of larval metamorphosis and phenotypic disparity in coral reef fish. Like marine invertebrates, the environment can also modulate different aspects of fish metamorphosis from pelagic larvae to reef-associated juveniles. Vincent presented a case of environmental acclimation in fish populations living in the enclosed lagoon of Taiaro Atoll, an extreme environment in French Polynesia, (Reynaud et al. 2023). Compared to their natural habitat (beaches and mangroves), the lagoon displays higher salinity and temperature. Interestingly, fish living in the lagoon and their counterparts living in oceanic reefs were identical at the genetic level, despite differences in their habitat. To understand the molecular mechanisms of fish acclimation to high salinity, Vincent and colleagues compared their transcriptomic profiles and found that each population had particular differences in thyroid hormone regulation. Thyroid hormones are central hormone regulators triggering metamorphosis in vertebrates (Zwahlen et al. 2024). In fish, they regulate a major metabolic shift during metamorphosis from the juvenile to the adult phase, including changes in pigmentation and maturation of the nervous/sensory and digestive systems. Vincent and colleagues proposed a new view of the role of thyroid hormones: rather than functioning independently in metamorphosis, metabolism and seasonality, thyroid hormones perform a 3-in-1 function of timing a biphasic life cycle transition. Thus, local acclimation to the environment is an endocrine-driven process, conferring plasticity on fish from the time they undergo metamorphosis and enabling them to adapt to high salinity conditions.
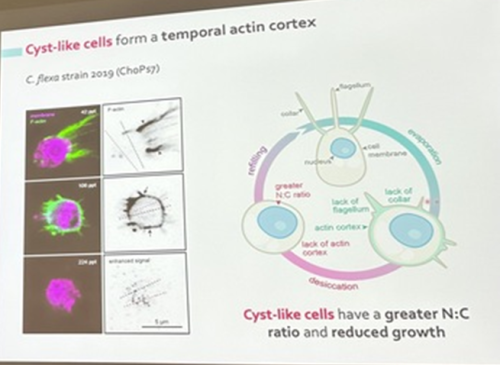
Núria Ros-Rocher, postdoctoral researcher at Institute Pasteur (Paris, France), presented another case of salinity-dependent life cycle transition in a microeukaryote. Núria showed that Choanoeca flexa, a facultatively multicellular choanoflagellate (Brunet et al. 2019;Reyes-Rivera et al. 2022), develops colonies by an unconventional mixed mode of clonal-aggregative multicellularity (Ros-Rocher and Reyes-Rivera et al. 2024). To further explore the ecological determinants of C. flexa clonal-aggregative multicellularity, Núria, Josean, and colleagues combined cell biology approaches in the laboratory with fieldwork experiments in the Caribbean island of Curaçao, the original site where this species was discovered (see Brunet et al. 2019). In the field, they surveyed 150 seawater-filled splash pools, the natural habitat of C. flexa, and explored the impact of splash pool evaporation and refilling cycles on C. flexa multicellularity. C. flexa colonies could only be observed in a narrow range of salinity and were never observed in higher salinities in the field, yet sometimes appeared in splash pools that got refilled by waves or splash after complete desiccation.
Surprisingly, multicellular C. flexa colonies could be recovered from soil samples collected from dry splash pools upon rehydration with seawater in the lab. Later experiments in the lab mimicking the evaporation of splash pools showed that colonies dissociate into quiescent single cells in high salinities, losing multicellularity. In those conditions, some single cells transitioned into a desiccation-resistant form, losing some of the stereotypical choanoflagellate features (a collar and a flagellum). Repeating similar rehydration experiments, Núria showed that those cells could return to their multicellular form upon rehydration. The authors hypothesized that this mixed clonal-aggregative strategy might represent a versatile adaptation to the ephemeral nature of splash pools where the simultaneous action of both mechanisms could allow for a faster re-establishment of multicellularity. In ongoing work, Núria, Josean and colleagues are exploring the genetic diversity of newly isolated strains from distinct splash pools. They are focusing on kin recognition capacities that may impact choanoflagellate survival in the field and provide insights into the adaptation of this organism in a highly fluctuating environment.
Núria Ros-Rocher identified a cyst-like stage in a choanoflagellate life cycle
In addition to metamorphoses and multicellularity, three presenters also discussed later-in-life transitions related to reproduction. Mariella Ferrante, researcher at the Stazione Zoologica Anton Dohrn (Italy), presented recent work on centric and pennate diatoms exploring how these different single-celled microalgae switch between sexual and asexual reproduction. While sexual reproduction in centric diatoms can be triggered by environmental cues such as light and salinity, pennate diatoms do not appear to have a specific trigger for initiating sexual reproduction. However, planktonic pennate diatoms need to solve the challenging problem of locating a mate within the vast 3D ocean to undergo sexual reproduction before they drop below a critical size due to several rounds of asexual reproduction which cause them to decrease in size with each asexual division. Mariella and colleagues are using RNA-Seq and single-cell RNA-seq in different diatom species and reproductive stages to explore heterogeneity in diatom populations as well as to identify a panel of molecular markers for detecting diatom sex events at sea (Annunziata et al. 2022).
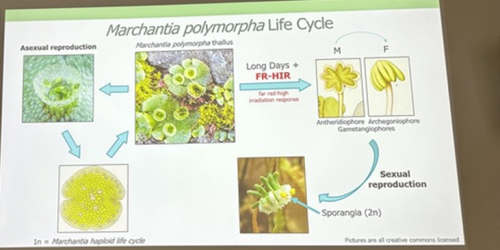
Alexandre Alié, researcher at the Laboratoire de Biologie du Développement (LBDV) in Villefranche-sur-Mer, is also using scRNA-Seq to understand asexual reproductive transitions in another organism that inhabits the pelagic oceans – Mediterranean salps, planktonic tunicates that form large chains through asexual reproduction. Alexandre and colleagues hope to use this information to understand more about how the stem cells in the stolons of the salps can bud off dozens of new individuals and how this process is influenced by environmental factors such as temperature and food availability. Finally, David Hoey introduced us to the molecular regulation of transformations for reproduction in the liverwort Marchantia polymorpha, which switches between sexual and asexual reproduction throughout the year depending on light quality, day length and drought conditions (Duckett et al. 2024). David generated knockout lines of a RAS gene and found that it is a regulator of asexual reproductive development in M. polymorpha. Remarkably, this gene is responsive to both light and drought cues.
David Hoey introduced us to the life cycle of the liverwort plant, Marchantia polymorpha
Overall, a strong theme running through the symposium was the development of molecular lab techniques such as gene knockout and single-cell RNA-Seq for application in a variety of non-model organisms. Ongoing studies on all of these emerging models are advancing our understanding of how environmental cues are detected and lead to important transitions in a life cycle. We also found inspiration in the examples from several researchers of combining their molecular lab studies with field studies of their organism of interest, to place our understanding of organismal and life cycle diversity back into a broader ecological context. We anticipate that this will be a strong theme going forwards, and aligns with the emphasis on eco-evo-devo that was highlighted at this year’s conference by keynote speaker Scott Gilbert, who also drew attention to the importance of microbial ecology in shaping animal life cycles and symbioses. Could inter-species or inter-kingdom interactions be a future focal area to further inform life cycle evolution?
References
Annunziata et al. 2022. Trade-off between sex and growth in diatoms: Molecular mechanisms and demographic implications. Science Advances 8:3, DOI: 10.1126/sciadv.abj9466
Campli et al. 2024. The moulting arthropod: a complete genetic toolkit review. Biological Reviews of the Cambridge Philosophical Society, https://doi.org/10.1111/brv.13123
Duckett et al. 2024 The biology of Marchantia polymorpha subsp. ruderalis Bischl. & Boissel. Dub in nature. Frontiers in Plant Science, 15, https://doi.org/10.3389/fpls.2024.1339832
Hird et al. 2024. Microalgal biofilm induces larval settlement in the model marine worm Platynereis dumerilii. Royal Society Open Science, https://doi.org/10.1098/rsos.240274
Nakanishi et al. 2015. Sensory flask cells in sponge larvae regulate metamorphosis via calcium signaling. Integrative and Comparative Biology 55(6): 1018-1027, DOI: 10.1093/icb/icv014
Reynaud et al. 2023. The environment: A vector of phenotypic disparity during the settlement phase of coral reef fishes. Journal of Experimental Marine Biology and Ecology, 568: 151937, https://doi.org/10.1016/j.jembe.2023.151937
Reyes-Rivera et al. 2022. Nitric oxide signaling controls collective contractions in a colonial choanoflagellate. Current Biology, 32(11): P2539-2547, DOI: 10.1016/j.cub.2022.04.017
Ros-Rocher et al. 2024. Mixed clonal-aggregative multicellularity entrained by extreme salinity fluctuations in a close relative of animals. bioRxiv, https://doi.org/10.1101/2024.03.25.586565
Zwhahlen et al. 2024. The ecological function of thyroid hormones. Philosophical Transactions of the Royal Society B, Biological Sciences, https://doi.org/10.1098/rstb.2022.0511
Acknowledgements
The Company of Biologists provided financial support for this symposium. We thank the organisers of the EED2024 conference in Helsinki, Finland, and all attendees of this symposium for contributing to discussions of this session. We also thank the symposium speakers for contributing to revision of this report.
Given all the outstanding evo-devo work going on in the London area, we feel that a regular gathering to share recent discoveries and to network would be a real asset. Not too long ago there was such a meeting, and we are aiming to revive the spirit of it.
Our hope is to arrange a sustainable meeting that would occur once or twice a year. The current plan is to have half-day events that include short presentations/feedback (15 min + 5 min Qs) and socialization. Ideally, the location of these meetings will rotate among the organizer’s campuses. We wish to cast a wide net by defining “evo-devo” broadly and attract researchers working directly in the area but also those working on evolutionary or developmental biology looking to bridge to the other side.
We have arranged the rebooting of London EvoDevo to be November 8th on Queen Mary’s campus from 13:00 – 17:00. We hope that you or people in your group will join and potentially be willing to either apply for a talk for this meeting or the future ones.
For more information and to register, please follow this LINK.
We hope to see you in November!
London EvoDevo Organizers,
Ferdi Marlétaz (UCL)
Margarida Cardoso Moreira (Crick), James DiFrisco (Crick),
Chema Martin (QMUL), Alex de Mendoza (QMUL), Roman Arguello (QMUL)
In this SciArt profile, we meet Yoshinari Yoshida, a philosopher of science focusing on the conceptual and historical issues in developmental biology. Yoshinari enjoys drawing animals and plants, and have recently got into drawing primates, such as gibbons and orangutans, because of his research into in vitro models in comparative biology.
Can you tell us about your background and what you work on now?
I was born in Milton Keynes, UK, and grew up in Japan. I did my undergrad and Master’s in Philosophy and History of Science at Kyoto University, Japan, and Ph.D. in Philosophy at the University of Minnesota, USA. I am currently a postdoctoral fellow at the University of Exeter, UK. Throughout this academic career, I have studied philosophy of science, with a special focus on conceptual and historical issues in developmental biology. My central philosophical interest is in how various models and representations (e.g., “living models” such as model organisms and cell culture systems, and visual models such as diagrams and illustrations) play their roles in the production of knowledge about generality and diversity in developmental processes. My current project focuses on the prospects and limitations of in vitro models in comparative biology. I try to clarify how in vitro systems, such as organoids, play their roles as models in the context of cross-species comparison and reconstruction of evolutionary history. I examine experimental and inferential practices as well as interdisciplinary interactions among experts to tackle this philosophical question.
Catalina the Cat – digital illustration.
Were you always going to be a philosopher of science?
When I was younger, I was rather going to be a scientist. I was a kid who loved insects and extinct animals, and in my teenage era, I enjoyed learning about evolution and embryonic development. But I also liked the philosophical discussions about conceptual questions. When I was in the second year of my undergrad, I took the courses “philosophy of science” and “history of science.” I enjoyed these courses so much, and started thinking that studying these fields would allow me to fulfill both my interest in science and my love of philosophical discussions. I decided to major in philosophy of science, and since then, I’ve been in this field.
Brachiating gibbon – digital illustration. Gibbons are my favorite primates. I’m impressed by how efficient their locomotion is! I used a photo by shilmar in Pixabay as a reference.
And what about art – have you always enjoyed it?
Almost yes. I was already into art when I was very little. I was a kid who kept drawing during lunch break in elementary school as well as after school. My favorite objects were monsters, robots, and manga characters. There were periods in my life when I was a bit away from doing art, such as during high school. But I’ve maintained my interest in art. Several years ago, when I was in my Ph.D., I bought a tablet and started drawing digitally. Until then, drawing had been a private hobby just for myself. But I began posting my artwork on social media and also drawing more constantly.
Sitting gibbon – digital illustration. I used a photo by Kadisha in Pixabay as a reference.
What or who are your most important artistic influences?
It’s not easy to name one specific artist. (I still lack systematic knowledge of art history.) But I’m often inspired by realistic oil paintings from the modern and early modern periods. My favorite is those paintings that are so realistic that they look as if they are photographs from distance, but consist of explicit strokes when seen closely. Those paintings make me think about how we see the world.
Orangutan – digital illustration. It was really fun to draw the unique three-dimensional shape of the face. I used a photo by Hollasue in Pixabay as a reference.
How do you make your art?
Currently I do everything using a painting app called Procreate in my tablet. When I see an object or scene whose shapes, colors, textures, or patterns I find interesting, I take pictures, which I may use as a reference later. Or, sometimes I become interested in a certain animal or plant, and search for images of the animal or plant and then start drawing it.
Radishes – digital illustration.
Does your research influence your art at all (or vice versa), or are they separate worlds?
Art is my major hobby that is a source of joy distinct from research in my life. But I think there are some connections between the two. For instance, I am interested in the role of images in science. I have some articles (one published, one currently under review) about how the use of specific types of diagrammatic representations contribute to knowledge production in science. I am also working on a side project with my colleagues about the role of speculation in evolutionary thinking, in which we discuss the power of illustrations. I think this philosophical interest of mine in images in science has a common root with my artistic interest in drawing and painting. My art is also sometimes influenced by my academic interests. For example, I’ve recently been into drawing primates, such as gibbons and orangutans. This was triggered by my current research project on in vitro models in comparative biology. I’ve read studies that apply advanced in vitro technologies to primates to elucidate human evolution, which made me interested in primates themselves.
Puffin – digital illustration. I used a photo by mystraysoul in Pixabay as a reference.
What are you thinking of working on next?
There are a few ideas. I want to keep working on primates and drawing as many primate species as possible. That would become an interesting series! I’m also interested in making art that is related to developmental biology. Even though my primary philosophical focus has always been on developmental biology, I’ve not made art about this field. It would be fun to make drawings inspired by the normal developmental stages of chick embryos, mechanisms of collective cell migration, or tissue movements in gastrulation. Finally, I’m interested in trying oil painting. While I’ve drawn only digitally over the last several years, I love seeing oil paintings at museums. That would be an interesting new step.
In our recent paper published in Developmental Biology entitled “Localization and origins of juvenile skeletogenic cells in the sea urchin Lytechinus pictus”, we studied the intriguing and unique late larval development of sea urchins with a focus on skeletogenic cells, which deposit the biomineral skeleton. Here, I discuss how this project came about, why we were excited by our findings, some of the challenges faced, and what’s next for the project.
Project beginnings
In 2021, I joined Deirdre Lyons’ lab at Scripps Institution of Oceanography (UC San Diego) to explore my fascination for invertebrate evolution and development. I was excited to develop a project on skeletogenesis and biomineralization in sea urchins, a topic in which Deirdre and I shared interest. As I was new to the field, I dove into the literature and realized that although I was interested in embryonic development, I was most fascinated by the development of the adult body plan, which occurs throughout the late larval stage. This study was also motivated by the development of the sea urchin Lytechinus pictus as a better model species [1-3], which has been an ongoing collaboration between the Lyons lab and the Hamdoun lab, also at UC San Diego. Since L. pictus is a faster developing and more transparent species than other commonly studied sea urchins, we could study and image late larval development in higher detail.
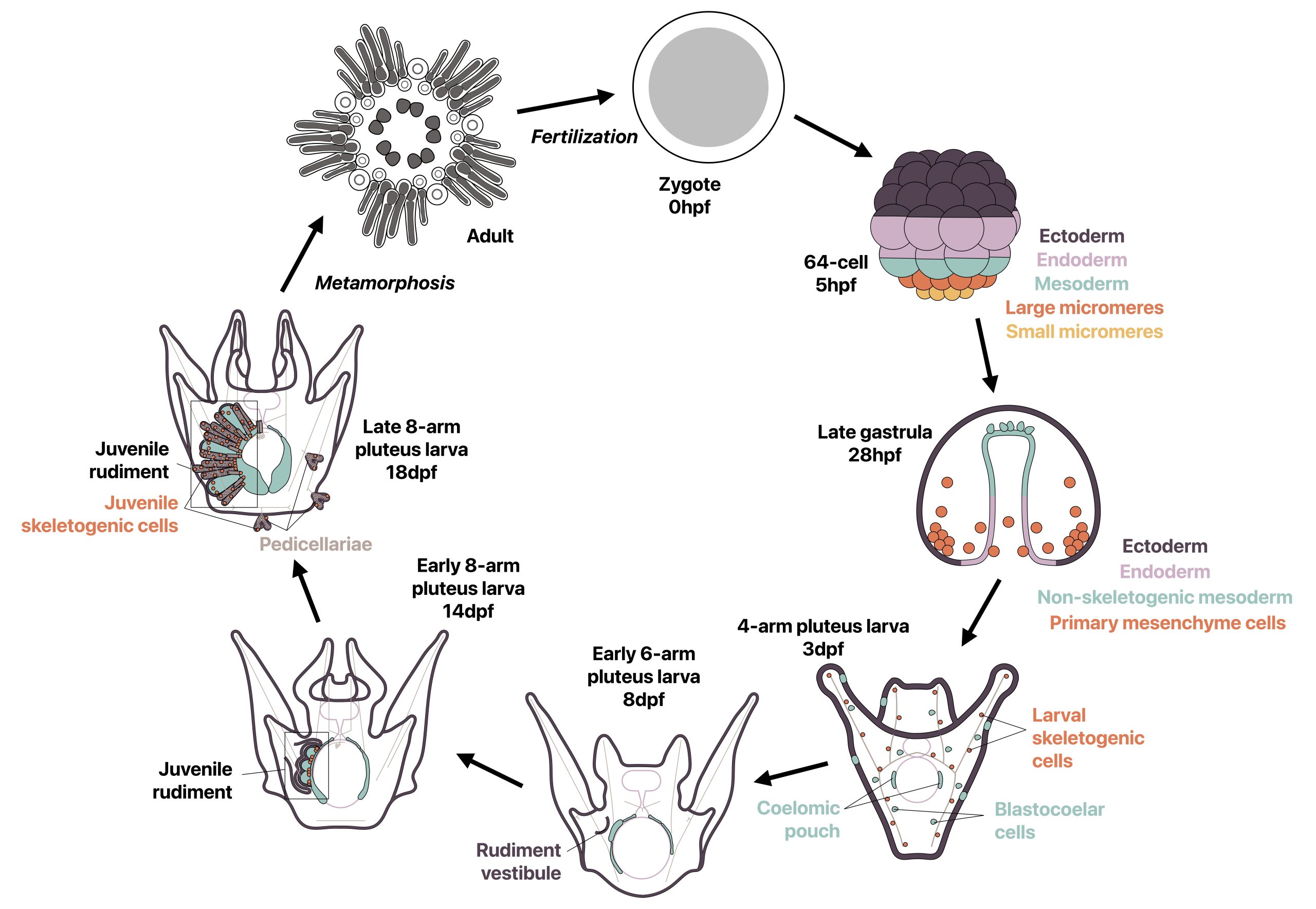
Most sea urchins are indirect developers and undergo a complete switch in body plan during metamorphosis from a bilaterally symmetric swimming larva to a pentaradially symmetric benthic juvenile. Prior to metamorphosis, much of the adult body plan develops on the left side of the larva in a structure called the rudiment, while other smaller adult structures develop in other locations (Fig. 1). I wondered how two drastically different body plans could be made from the same set of genetic information, and how the development of a second body plan could occur outside of a classic embryonic context. I was specifically inspired by a study from 2007 by Mamiko Yajima in which she demonstrated that the juvenile skeletogenic cells arise not from the larval skeletogenic lineage, but from mesodermal cells that had a non-skeletogenic origin in the larva [4], and I found it fascinating that cells with one fate in the larva can be completely reprogrammed to make a new cell type in the adult. I therefore decided to revisit this topic using recent technical advances in the field.
Prior to this study, rudiment development had been described in detail at the tissue level and some immunostaining had been performed [5-9], however mRNA localization via in situ hybridization was challenging due to the thickness and complexity of the late larval tissue. By leveraging HCR RNA-FISH, we were able to conduct some of the first mRNA in situ hybridizations in these late stages as the small probes used in this technique are capable of penetrating into thick late larval tissue. I chose to focus on skeletogenic cells as they have been well-studied during embryonic development of sea urchins for gene regulatory networks and biomineralization [10,11] and had previously been studied in some detail in the late larva [7,8], although not in the newer model species L. pictus. We therefore asked three main questions: where are skeletogenic cells and biomineral located during late larval development in L. pictus within the complex rudiment morphology, what are the most likely origins of the juvenile skeletogenic cells, and are gene expression patterns across skeletogenic cells similar in embryonic and juvenile skeletal development?
Figure 1: Development of the sea urchin Lytechinus pictus. Most of the juvenile body plan develops prior to metamorphosis in the rudiment on the left side of the larva, which begins developing at 8 dpf. Metamorphosis occurs when rudiment is fully developed at 18-21 dpf.
What we learned
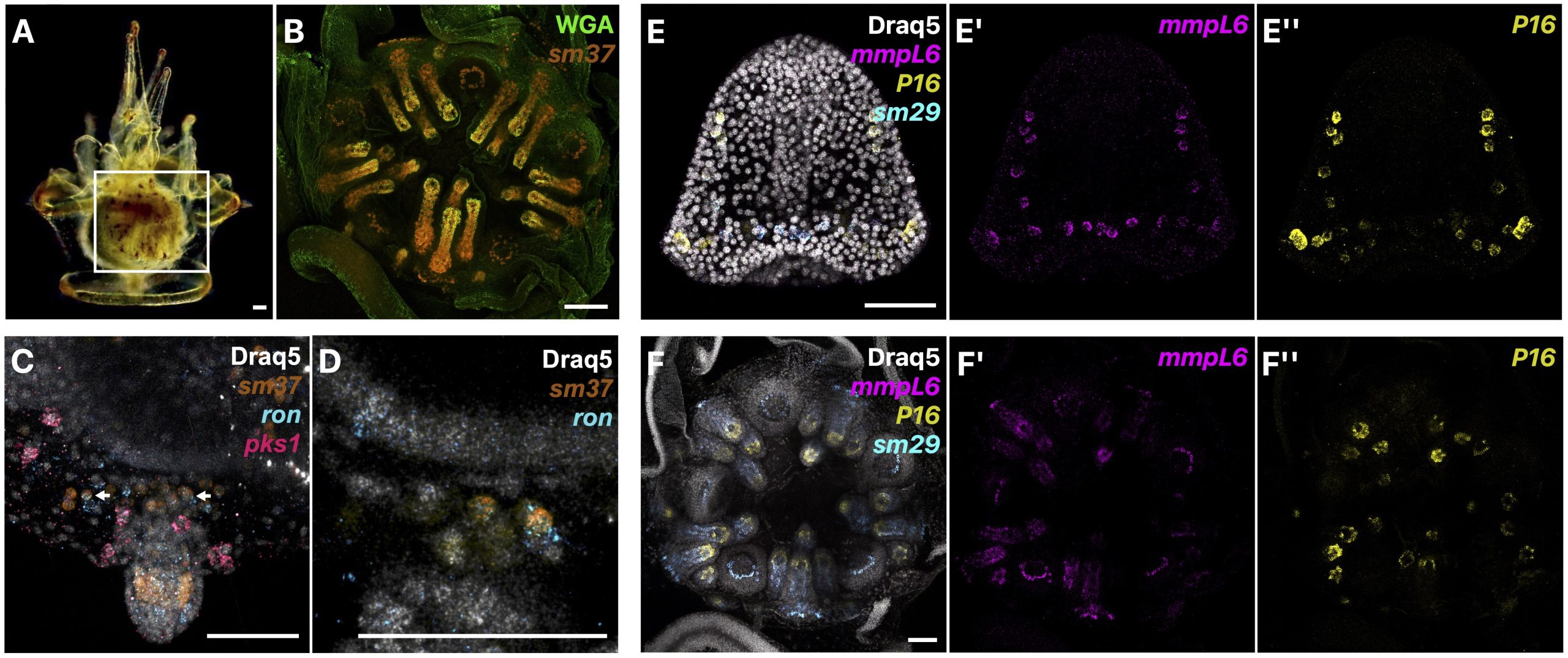
We first described in detail the localization of presumptive juvenile skeletogenic cells prior to biomineralization of juvenile skeleton, and at later stages, the localization of biomineral and juvenile skeletogenic cells both within and outside of the rudiment in L. pictus (Fig. 2A-B). We were excited to find that prior to the start of juvenile skeletogenesis in the rudiment, we could identify populations of skeletogenic cells using known biomineral marker genes that were not part of larval rods. Additionally, in some of these presumptive juvenile skeletogenic cells, we found co-expression of our skeletogenic cell marker and a marker of blastocoelar cells, which in the larva act as migratory immune cells (Fig. 2C-D). We therefore proposed that since mesodermal cells of non-skeletogenic origin contribute to juvenile skeletogenic cells [4], that blastocoelar cells may at least in part be the cell type that switches to a skeletogenic fate. We then sampled various biomineralization genes from different gene families and found that mRNA expression patterns throughout juvenile skeletogenesis are similar to the patterns during embryonic development (Fig. 2E-F).
Overall, our work gives insight into how indirect developers, like the sea urchin, can reuse cells and developmental mechanisms to make a new body plan from another. Additionally, it is thought that since all echinoderms have adult skeleton but not all have larval skeleton, that the larval skeleton in sea urchins may be a more recent innovation, while the adult skeletogenic cells could arise from a common origin [12]. Since other echinoderms have blastocoelar-like lineages in the larva [13-14], it is possible that this cell type is the ancestral adult skeletogenic cell lineage that expresses an ancestral skeletogenic gene regulatory network, although this hypothesis needs to be clarified with further work.
Figure 2: A) Late larva at 18 dpf in the left view with a box highlighting the location of the rudiment. B) Close-up of the rudiment at 18 dpf with HCR RNA-FISH for a biomineralization gene (sm37) and WGA stain to highlight biomineral. C-D) Co-expression of a blastocoelar (ron) and skeletogenic marker (sm37) in some cells around juvenile structures. E-F) Examples of skeletogenic gene expression patterns during embryogenesis at 32 hpf (E) and during adult body plan development in the rudiment at 18 dpf (F).
Unexpected challenges
One of the most challenging aspects of this project was identifying a consistent staging scheme during late larval development, which was an important part of the project to help establish L. pictus as a model sea urchin. Interestingly, the larval body plan and parts of the juvenile body plan, like the rudiment, often develop asynchronously, and so similar looking larvae may not have a rudiment at the same developmental stage [5-9]. Additionally, the extent of rudiment development was often different when metamorphosis occurred, leading to post-metamorphic juveniles in different developmental stages. Although frustrating when trying to stage the animals, this helped me think about development in new ways since in indirect developers, adult body plan development may not be as clear-cut as in embryogenesis.
I was also personally challenged to learn about all aspects of the scientific process, from asking clear scientific questions, thinking hard about the best ways to answer them, and learning to tell a compelling story through publication. As with any scientific project, there were many challenges and failed experiments along the way, particularly as I was learning many techniques for the first time. For instance, learning how to mount, orient, and image these large samples added a lot of time to the project, and I often had to retake very long acquisitions when images did not turn out as expected. This really challenged my ability to persevere when experiments were not working, but I felt especially honored and reinvigorated when one of our images was featured on the cover of Developmental Biology. Overall, I was very proud to have finished this work within the relatively short timeline of my master’s degree, and I am greatly looking forward to applying this newfound knowledge in future work.
What’s next with the sea urchins?
With this paper, we highlighted that sea urchin adult body plan development is fascinating, yet understudied, and how it is important to think of body plan development outside of an embryonic context. However, there are still many remaining questions to be answered. For instance, only skeletogenic cells have been studied in some detail during adult body plan development, and HCR RNA-FISH provides the opportunity to understand whole body patterning by investigating other cell types. However, more complex questions, such as mapping where adult cells are originating, would require more complex techniques. With the development of L. pictus as a better system where mature adults can be raised from egg in 4-6 months and with which transgenic lines can be created [1-3], an abundance of new questions could be studied. I am greatly looking forward to continuing work with this amazing animal in the future to answer innovative questions in adult body plan development and evolution. Finally, I am very thankful for my mentors, particularly Dr. Lyons, for the mentorship throughout this project, as well as other collaborators and colleagues who have supported and guided me through past and present research.
Figure 3: Video of a sea urchin mid-metamorphosis, with the juvenile body beneath the slowly metamorphosing larval body.
References
Nesbit KT, Fleming T, Batzel G, Pouv A, Rosenblatt HD, Pace DA, Hamdoun A, Lyons DC. 2019. The painted sea urchin, Lytechinus pictus, as a genetically-enabled developmental model. Methods in Cell Biology. 150: 105-123.
Jackson EW, Romero E, Kling S, Lee Y, Tjeerdema E, Hamdoun A. 2024. Stable germline transgenesis using the Minos Tc1/mariner element in the sea urchin Lytechinus pictus. Development. 151(20).
Vyas H, Schrankel CS, Espinoza JA, Mitchell KL, Nesbit KT, Jackson E, Chang N, Lee Y, Warner J, Reitzel A, Lyons DC, Hamdoun A. 2022. Generation of a homozygous mutant drug transporter (ABCB1) knockout line in the sea urchin Lytechinus pictus. Development. 149(11).
Yajima M. 2007. A switch in the cellular basis of skeletogenesis in late-stage sea urchin larvae. Developmental Biology. 307(2): 272-281.
Cameron RA, Hinegardner RT. 1978. Early events in sea urchin metamorphosis, description, and analysis. Journal of Morphology. 157(1): 21-31.
Heyland A, Hodin J. 2014. A detailed staging scheme for late larval development in Strongylocentrotus purpuratus focused on readily-visible juvenile structures within the rudiment. BMC Developmental Biology. 14(22): 1-14.
Yajima M, Kiyomoto M. 2006. Study of larval and adult skeletogenic cells in developing sea urchin larvae. The Biological Bulletin. 211(2): 183-192.
Smith MM, Smith LC, Cameron RA, Urry LA. 2008. The larval stages of the sea urchin, Strongylocentrotus purpuratus. Journal of Morphology. 269(6): 713-733.
Formery L, Wakefield A, Gesson M, Toisoul L, Lhomond G, Gilletta L, Lasbleiz R, Schubert M, Croce JC. 2022. Developmental atlas of the indirect-developing sea urchin Paracentrotus lividus: From fertilization to juvenile stages. Frontiers in Cell and Developmental Biology. 10.
Davidson EH, Rast JP, Oliveri P, Ransick A, Calestani C, Yuh C-H, Minokawa T, Amore G, Hinman V, Arenas-Mena C. 2002. A Provisional Regulatory Gene Network for Specification of Endomesoderm in the Sea Urchin Embryo. Developmental Biology. 246(1): 162-190.
Wilt FH. 2002. Biomineralization of the Spicules of Sea Urchin Embryos. Zoological Science. 19(3): 253-261.
Koga H, Morino Y, Wada H. 2014. The echinoderm larval skeleton as a possible model system for experimental evolutionary biology. Genesis. 52(3): 186-192.
Tamboline CR, Burke RD. 1992. Secondary mesenchyme of the sea urchin embryo: Ontogeny of blastocoelar cells. Journal of Experimental Biology. 262(1): 51-60.
Metchnikoff E. 1893. Lectures on the Comparative Physiology of Inflammation: Delivered at the Pasteur Institute in 1891.
The definition of outreach is “to reach further than”, however, I have found that when it comes to science outreach initiatives, we are only reaching those already in the loop. Although I personally applaud all these events and ideas and have been part of them for many years – either recruiting to my graduate program or showcasing my institution – it always seems like I am speaking to people who are already familiar with what a PhD program is or what academic research entails, and they typically seem to know which career or degree they want to pursue. I believe the key is to approach and interact with those who are not part of the scientific community.
As an international student, I thought that if I wanted to pursue a career in any way related to healthcare, the only road was medical school. It was not until pretty far into high school that I learned of alternate careers and graduate school. Maybe I am the anomaly, but I would bet that many students in similar positions do not know the many opportunities that are available by pursuing a graduate degree in biomedical sciences. Even though I have now moved on from my graduate training, I wish to remain involved in outreach initiatives. However, I want to do so a little differently, and I hope I can encourage others to do the same.
My idea of outreach will now not be limited to going to a local college to speak about my graduate program and such. I want “to reach further than” I have previously and introduce this amazing career path to people even before they reach high school, to truly go out on a limb and pitch this life to those that are not aware of it,rather than simply preaching to the choir. If we were to ask the average 3rd grader to describe a scientist, I’m sure they would all describe the typical Albert Einstein-esque look – messy hair, old and a little unhinged. And (although we all know someone like this in academia) I want to be part of outreach initiatives that challenge this idea, and instead pitch a lifestyle of endless curiosity and non-stop learning to kids that do nothing but wonder and ask questions themselves. If you had told me when I was a kid myself that I would be working with the technology used in Jurassic Park (CRISPR, gene editing, in-vitro cultures), I would have thought that it was the perfect job. I believe I can sell this same idea to kids now: kids that do not know what a PhD is, that do not think being a scientist is something that is feasible for them, or that have not met someone who is as passionate for learning as they are even later in life. There is a reason why younger kids are better at learning new languages; they are just ready to absorb all the knowledge you throw at them. So, this is the time to go and show them a science experiment, hook them in, and get them to ask “why?”.
I’m going to end my rant here, but I just think there is more we could do, there is more I want to do. Institutions around the world are starting initiatives specifically aimed at primary school children such as The Crick’s annual Discovery Day, or the Vanderbilt Institute for Infection, Immunology and Inflammation ‘MEGAMicrobe’ events which target students ages 5 to 14. This is a great starting point, and I hope that more initiatives like these start to take off, and more people are encouraged to get involved with them. Outreach should not be exclusive to the people already in the loop, the point of it all is to go beyond, and “to reach further than”.
What is one outreach event you have been involved in recently? What ideas do you have or have seen that could be used as science outreach? Comment below!
This article is co-written by Eliza Hirsch and Paul Bump.
Who are we?
Hi, my name is Eliza Hirsch, and I am an undergraduate in the Srivastava Lab at Harvard University. And my name is Paul Bump, and I am a postdoctoral fellow at the Srivastava Lab who works with Eliza. Our lab studies the highly regenerative acoel, Hofsteniamiamia. This Hofstenia species originally was collected in Bermuda and now lives in Cambridge, Massachusetts where we study stem cell biology and the evolution of regeneration across animals.
Who is Hofstenia?
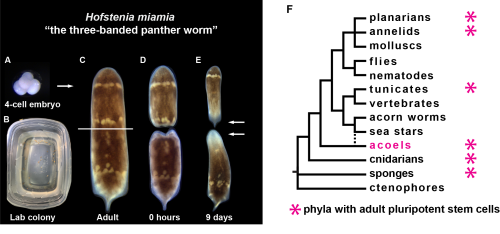
Hofstenia miamia is an acoel worm capable of whole-body regeneration, meaning that if you were to bisect one, both halves would regenerate, leaving you with two worms instead of one. Hofstenia maintain a population of adult pluripotent stem cells (aPSCs) that are responsible for regeneration. The regenerative ability of Hofstenia is quite impressive but they are hardly the only systems in which such regenerative ability exists. So why study them? Well, two other key traits of Hofstenia are central to answering this question.
Hofstenia lay embryos (A) in the lab in Tupperware (B) which develop into highly regenerative worms (C) that, upon amputation (D), can generate new heads and new tails in less than ten days (E) (white arrows). Hofstenia belongs to a group of animals called acoels (F), one of the many groups of animals with adult pluripotent stem cells.
Hofstenia have experimentally accessible embryos, something that is very central to the work we do on the development of the aPSCs. They lay their embryos when they are zygotes, allowing us to study their development from the single cell stage all the way up through hatching and beyond. Hofstenia embryos are also easily accessible due to their abundance. The worms we care for lay zygotes frequently enough and at a high enough volume that we can collect almost every day of the week and on most days, we get a substantial number of them. Thus, as a system Hofstenia allow us to study the development of a highly regenerative organism as well as its adult regenerative ability.
How to work with Hofstenia
To understand what it is like to work daily on Hofstenia we thought it would be best to give a perspective from Eliza who is completing her undergraduate thesis research in the Srivastava Lab:
Hofstenia are relatively easy to care for. In the lab, we keep a large portion of our worm colony in plastic Tupperwares of various sizes with up to sixty worms being stored in any given large Tupperware box. When not being cleaned or collected from or used for any other experimental processes, these boxes sit in incubators. The worms themselves live in artificial sea water that is made in the lab, something which perhaps requires the most time and effort in the caretaking of these animals. Adult worms are fed recently hatched brine shrimp as Hofstenia require live food. Hatchlings and sick worms or worms that may be left unattended over a long weekend without cleaning are fed rotifers as they pose a far smaller risk to the pH of the water and by extension the health of the worms in the event of their death.
To clean the worms and avoid problems with ammonia build up in the water and the like, twice a week we replace the water and wipe down the inside of the container. An important element of worm care is the collection of embryos. The animals lay their embryos on the sides and bottom of the container, and I gently scrape at the walls with a glass pipette to dislodge them and then collect them with the same pipette and deposit them in a dish full of sea water. On top of the wild type worms, the lab also has various lines of transgenic worms that are more sensitive than the wild type worms and much harder to replace and therefore require even more careful care.
Recently the lab has started using a water system for many of the wild type worms that significantly reduces the amount of time spent cleaning the worms as they have circulating water and are no longer in boxes but rather are in tanks. However, since my work depends on very early embryos, my worms are still in boxes so that I have access to those embryos which are more difficult to collect in the water system.
Paul and Eliza discuss care of Hofstenia in their slightly less tropical home of Cambridge, Massachusetts
How to crack an egg, starting development in Hofstenia
For my project, I have been collecting embryos every day. Hofstenia embryos are surrounded with a transparent chorion (their eggshell) that must be removed for much of the work that I do with them. Nearly daily I perform a chemical dechorionation on the single-cell zygotes I’ve sorted from the embryos I collected that morning. For this process I use a combination of chemicals and then mouth pipette embryos one by one into twenty-four well plates filled with a combination of artificial sea water and a small amount of antibiotics. Antibiotics are an essential addition as removing the chorion strips the embryos of a key defense against infection. I then raise these one-cell embryos up without a chorion so that I can do experiments at specific developmental stages that would be too sensitive to the dechorionation process.
Eliza works on removing the chorion of the Hofstenia embryo before beginning her experiments
The future for Hofstenia
Since Hofstenia is still a relatively new research organism, there are many possible questions about them to try to answer in one’s own research. In the future, hopefully many labs will continue to increase our overall understanding of Hofstenia and use that information to evaluate evolutionary and mechanistic questions of how the stem cell lineage develops and allows the regeneration of any missing body part. While one drawback of such a new model system is that many methods for working on the animals have yet to be developed, the future is bright as we will continue developing technologies with which to study Hofstenia development and regeneration.

 (No Ratings Yet)
(No Ratings Yet)


 (2 votes)
(2 votes)