Authors: Sandra Franco-Iborra (ASAPbio Community Lead), Pablo Ranea-Robles (Postdoctoral Fellow, Novo Nordisk Foundation Center for Basic Metabolic Research at the University of Copenhagen), Lonni Besancon (Assistant Professor, Linköping University) & Jonathon Alexis Coates (Assoicate Director, ASAPbio; jonny.coates@asapbio.org)
Scientific publishing is experiencing a reckoning; scientists are resigning across a wide range of editorial boards, the dominance of traditional publishers has been successfully challenged, open access has revolutionized publishers’ business models, there’s a lack of trust in science and preprints are on the rise. Preprints (manuscripts shared prior to journal-organised peer review) offer an opportunity to transform scholarly communication into a system that places science and society first in addition to alleviating many of the issues we currently face.
Brief history of preprints in the life sciences
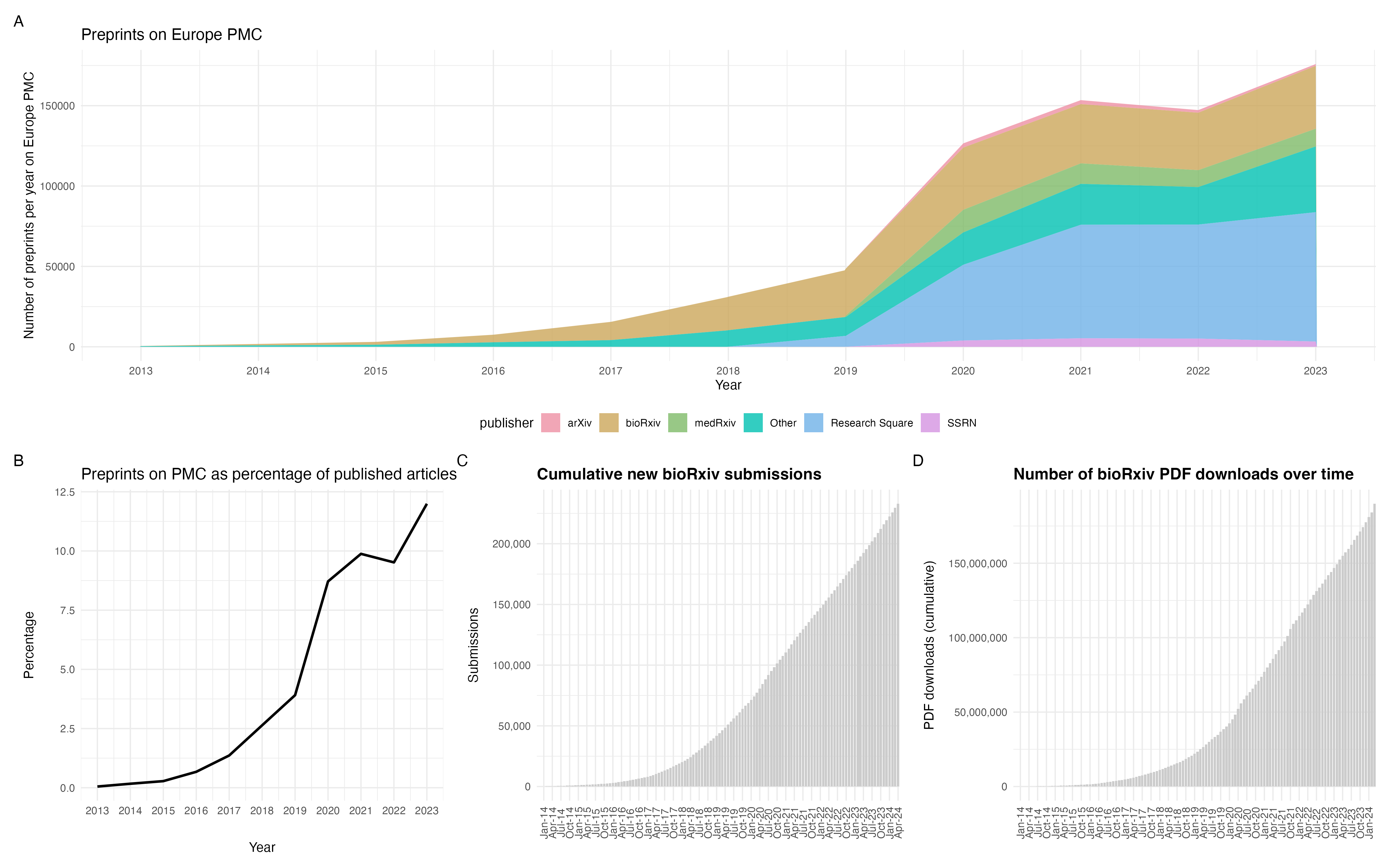
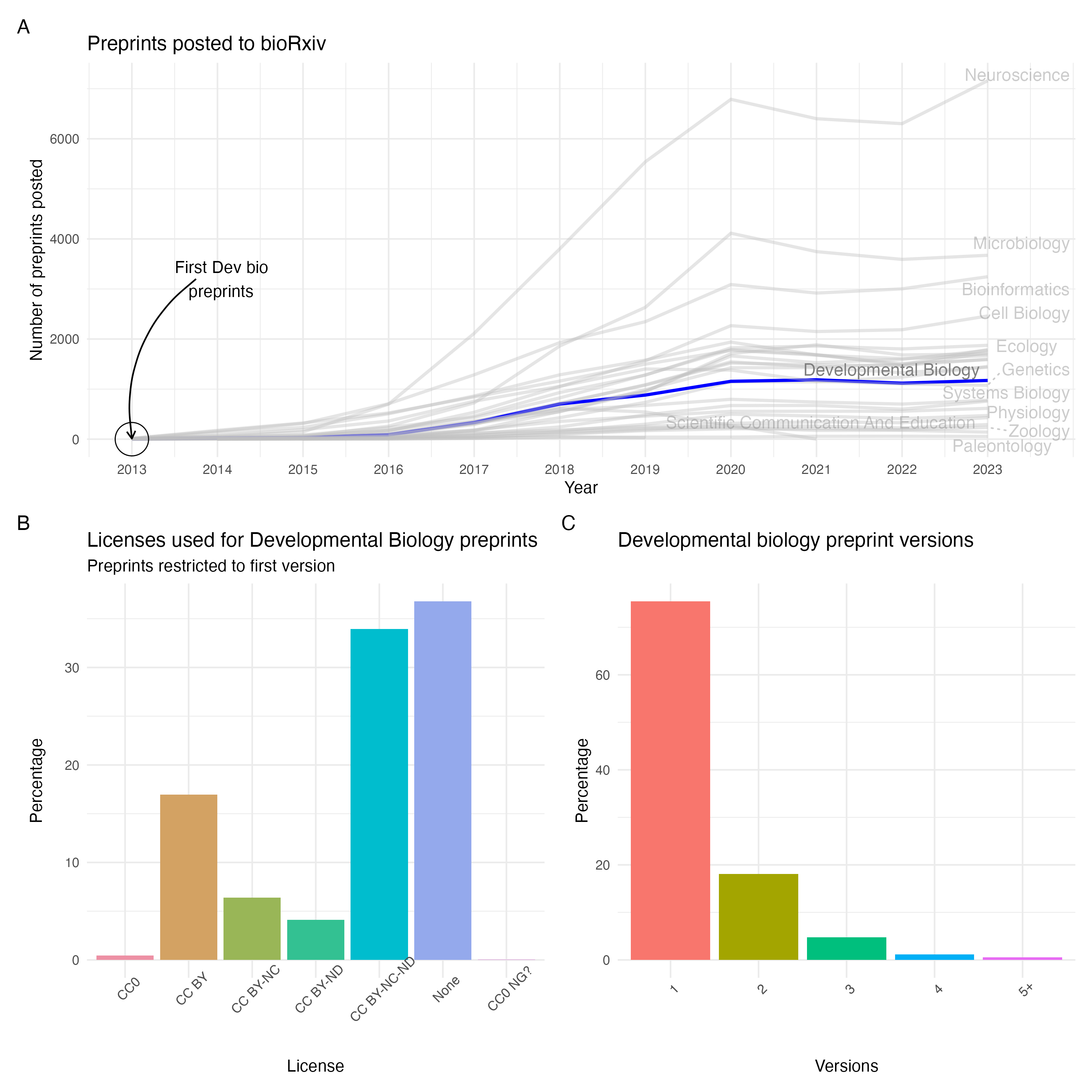
Although the fields of physics and mathematics are often credited as the origins of preprinting, the practice actually began in the 1960’s with biology focussed NIH information exchange groups (IEGs). However, this initiative faced opposition from publishers and scientific societies who effectively ended the IEGs by 1969. In 1991, arXiv launched and the physics community rapidly adopted preprinting. It was not until the launch of bioRxiv in 2013 that biology began to accept preprints. More recently, some funders such as the Chan Zuckerberg Initiative and Bill & Melinda Gates Foundation have begun to mandate preprints. We’ve also witnessed governmental declarations that support a not-for-profit, no cost to authors or readers system of academic publishing; a system that has preprints at the beating heart. There are now over 750,000 life science preprints (Fig 1A), representing approximately 12% of the literature in 2023 (Fig 1B). Of these 750,000, over 220,000 are hosted on bioRxiv alone (Fig 1C) and have been downloaded over 150 million times (Fig 1D).
Fig 1. Preprints in the life sciences. A) Preprints indexed on Europe PMC. B) Preprints indexed on Europe PMC as a percentage of published articles on Europe PMC. C) Cumulative bioRxiv submissions since 2013. D) Cumulative PDF downloads for bioRxiv preprints. Thanks to Europe PMC for the code and data to produce panels A & B and Nicholas Fraser for the code used to produce panels C & D.
Preprints in Developmental Biology
With the launch of bioRxiv in 2013, Developmental Biology was quick to adopt this new method of scientific communication, posting preprints that same year. As of 2023, Developmental Biology sits in the middle of preprints on bioRxiv, as the 15th largest field with almost 6700 total preprints posted (Fig 2A). This is also true in terms of preprint downloads where Developmental Biology preprints are downloaded slightly less than the mean for bioRxiv. Approximately ~70% of bioRxiv preprints are eventually published, with our data showing that at least 54% of Developmental Biology have been published by the end of 2023 (though this number is likely to be higher due to issues linking preprints to the published version). Additionally, many Developmental Biology preprints are posted with restrictive licenses with only 17% having an open, CC-BY license (Fig 2B). However, this is very similar to the larger bioRxiv corpus where only 18% of preprints have a CC-BY license. Similarly, most Developmental Biology preprints are posted as a single version which negates the benefit of being able to iteratively update a preprint (Fig 2C). Within the field, the University of Cambridge (UK) is one of the largest contributors of preprints, having contributed over 100 Developmental Biology preprints to bioRxiv since 2013.
Interestingly, despite being in the middle of the pack in terms of preprint posting, Developmental Biology is the second biggest category for number of preLights posted (>350). preLights (a preprint highlighting service from CoB) is a platform in which ECRs write news & views style articles of preprints. This highlights the benefits of a strong community-led curation and demonstrates how this can work for other platforms.
Fig 2. Characteristics of Developmental Biology. A) Developmental Biology preprints in bioRxiv over time. B) Licences used for Developmental Biology preprints on bioRxiv. C) Number of versions for Developmental Biology preprints on bioRxiv. Data downloaded using the bioRxiv API.
Develop your use of preprints
Preprints serve multiple purposes. First, they help to shift the power dynamic in academic publishing, placing the power back in the hands of authors. This helps to accelerate the dissemination of scientific knowledge, free from the pressures and delays associated with publishing. The benefits of this open, accelerated system were highlighted during the COVID-19 pandemic when almost 40% of the initial COVID-19 related research was first shared as preprints, directly leading to changes in policy and potentially saving lives. Preprints can appear online within 48 hours of submission, compared to months and years under traditional publishing routes. This also proves particularly useful to publicly rebut published research and avoid the long delays of scientific corrections or the rather unused platforms for post-publication peer review
Preprints also decouple the quality of research from metrics like journal impact factor, promoting fairer, better, evaluation. They also provide a platform for early-career researchers to showcase their work and gain recognition, fostering a more inclusive and diverse research landscape. Additionally, preprints can be used iteratively to share ongoing research and get feedback from the community. However, perhaps the most important benefit of preprints is that they offer a viable route towards meaningful change to scientific communication; one that is free from financial incentives and pressure and that is community-focussed.
The best way to get started with preprints is to use them in your research by citing and reading them. The Node maintains “In preprints” which is a great series for discovering relevant Developmental Biology preprints each month. Going further, you could also discuss preprints in your journal club – or even review them on platforms like PREreview or hypothes.is with support from ASAPbio. For ECRs, preLights is an excellent opportunity to take your first steps into commenting on preprints.
Ready to develop how you share your research, amplify your voice, and contribute to a more open, Developmental Biology community? Grab our checklist (Box 1), explore resources, and join the preprint movement!
As biologists we work regularly with images to collect, interpret and communicate our data, findings and ideas. These days, however, images are almost entirely digital, and it is becoming increasingly uncommon to incorporate manually drawn pictures or 3D hand-crafted models into research, either during experimental observations or to communicate findings. Accurate scientific illustration is an important skill to record anatomical detail during an organism’s life, and realistic drawing was the main working method for early naturalists and anatomists, but drawing and modelling can also be used as a way to develop new ways of thinking about topics and processes from a different perspective1,2. We set out to explore how hand-drawing and 3D modelling could allow researchers today to engage with their research from new perspectives in a collaborative workshop guided by artist Dr Gemma Anderson-Tempini.
On the afternoons of Wednesday December 13th and Thursday December 14th 2023, we held a drawing workshop to bring together participants across three different research groups – (1) the ‘Molecular Marine Systems’ group of Dr Elizabeth Williams in the Faculty of Health and Life Sciences at the University of Exeter, (2) the ‘Micromotility’ group of Dr Kirsty Wan from the Living Systems Institute (LSI) at the University of Exeter, and (3) the ‘Algal Microbiome and Ecophysiology’ group of Dr Katherine Helliwell at the Marine Biological Association (MBA), Plymouth. Although focused on different research questions from different perspectives, the groups share an overlapping interest in understanding how microscopic organisms can sense, respond to, and move through their fluid environment.
Welcome to the workshop. We looked at planktonic larvae and discussed the potential of drawing: to infer or ‘draw hypotheses’, to ‘be like’ a biological process, to select salient information, to show and share understanding, and to constructively collaborate towards a processual view (Photo – Kirsty Wan).
Study organisms across the research groups include marine invertebrate larvae, microalgae, particularly diatoms, and protists. How these organisms transition between distinct phases of their life cycle in response to specific environmental cues is also of common interest across the different groups. We took inspiration from Maria Sibylla Merian, an entomologist, naturalist and illustrator (1647-1717) who was among the first to depict animal life cycles in the context of their specific environments at each different life phase3.
All research group members were invited to participate in the workshop and participants included group leaders, postdoctoral research fellows, graduate students and research technicians. Prior experience with drawing or art ranged from absolute beginner to confident regular practitioner. Workshop participants included scientists from both biology and mathematics backgrounds.
The first day of the workshop started with brief roundtable introductions and an overview of research topics by Liz and the practice of using drawing to represent dynamic biological processes by Gemma. As a drawing ‘warm-up’, we started with a group exercise on the ‘evolution of shape’, based on the drawing process developed by Gemma as part of her previous ‘Isomorphogenesis’ project, a drawing-based enquiry into the shared forms of animal, mineral and vegetable morphologies2.
Each participant was given a 3D object to make an observational drawing. This drawing was then passed to the next participant, who added a connected object with an alteration suggested by selecting an action word at random. We continued the process until everyone’s initial drawing was returned to them, and observed the evolution of our original shapes. This allowed everyone to start drawing without feeling inhibited by the need to draw something perfectly, or the lack of an idea about what to draw.
Examples from the ‘evolution of shape’ group drawing exercise. A trend emerged across the group in that various forms started to accumulate cilia.
After this exercise, each participant was asked to draw a depiction of their own current research project or scientific question, and explain what it is they wanted their drawing to show. Examples of topics discussed included trying to understand how a microscopic organism could coordinate multiple cilia for effective swimming, or finding a new way to depict an organism’s life cycle by using continuous line drawing to highlight the connectivity between different life phases and promote the idea of considering an organism as its whole life cycle.
To conclude the first day of the workshop, participants were then asked to complete a drawing exercise similar to the initial ‘evolution of shape’, in which we imagined our organism of interest as a 3D shape, using questions to guide and direct the drawing: ‘What developmental stage is your organism?’, ‘What kind of environment is your organism in?’, ‘What type of response does your organism show?’. These questions also related to our prior recommended reading on the experience of larvae in flow4. In this exercise participants were encouraged to generate a bold drawing that would take up all the space available on their A3 blank sheet.
The instructions for the final drawing exercise on Day 1 (left), with an example outcome drawing of ‘worm larva movement in light and flow’ demonstrating an attempt to fill all the space on the page (right).
We started the second afternoon of the workshop with a short visualization exercise followed by discussion, to help bring participants into the present, and focus on their research questions. Participants lay or sat in comfortable positions with their eyes closed while Gemma guided them on a journey in the plankton as a microscopic organism, slowly dropping down deeper and deeper into the sea in search of a place to settle down on the sea floor. The effect of this exercise on participants was fascinatingly variable, with responses ranging from finding the experience stressful, busy and complicated, with many organisms jostling for position in the plankton, and the added complications of moving in a big 3D space, with different types of flow that could take an organism anywhere, and the ever-present threat of predators from every side. Other participants found the experience relaxing, due the perceived reduction in the types of decisions and actions they could take as a microscopic member of the plankton – sink or swim? The overall effect was to bring the group closer together and focused on the shared topic of marine microorganisms’ development and navigation.
Following the visualization exercise, participants were offered a choice of activities including free-drawing with or without tracing paper, a paper folding origami activity, or the use of a circular maze template that could be converted from 2D circle into a 3D cone. These activities provided a basis from which participants could develop their own ideas about their research project or question. Examples of projects included using transparent layers to add information about environment and different life stage priorities to a coral life cycle, mapping the settlement journey of planktonic larvae through a circular maze, using the maze to demonstrate the carbon capture process during diatom sinking, or to develop an anatomical map of cell types in a marine larva. Origami structures were used to explore the life cycle of a marine worm, incorporating research goals into different sections, to explore environmental effects on marine larvae with changeable combinations of environmental factors, or to demonstrate biodiversity and morphology of diatom species.
Mapping the planktonic journey of larvae using a maze template (left), and developing an interactive marine invertebrate life cycle in 3D with origami (right).
Following the independent work, we ended the second day with presentations and discussion of each participant’s work in progress. Consensus across the group was that drawing provided a useful way of thinking about research. There was strong interest in finding ways to incorporate drawing more into our research papers or use it as a tool to start thinking about and discussing new projects by drawing what the results could look like, the experimental plan, or the overarching question. This workshop showed us that drawing can be used to stimulate discussion, think about research projects, generate new ideas, images and hypotheses/questions, promote lab group interactions and understanding and collaboration between different research communities. Common themes that emerged from our discussions were the usefulness of drawing and paper craft as a tool for teaching and communicating, and to remind us of the bigger picture and broader impacts of our research.
Overall, it was useful to have some templates designed by Gemma to work with, such as mazes, games, or origami structures, which helped those unfamiliar with creative work or processual drawing, as these provided an initial framework from which to develop ideas. Representing biological phenomena such as metamorphosis, behaviour and movement as a process is not easy, but through the workshop interesting ideas emerged regarding both ways to represent a process dynamically, as well as ways to think about the process itself. For example, one participant developed a carousel model or zoetrope with which to demonstrate marine larval behaviours in response to changes in oceanic pressure, while another developed a diatom ‘teaching wardrobe’ with interchangeable layers allowing the demonstration of a diatom’s response to different environmental conditions.
Planning a zoetrope to demonstrate planktonic larval behavioural responses in action (left), and development of a ‘teaching wardrobe’ to demonstrate environmental effects on diatoms (right).
New ways of thinking about life cycles also emerged, in particular the representation of a life cycle as a spiral instead of the classic circle. This idea has an interesting synergism with the recent reflections by Sarah and Scott Gilbert on the prevalence of spiral forms in nature, and the possibility of thinking about the animal holobiont as two interlocking spirals, one representing the microbiome and the other the animal – ‘Circles are complete and perfect; life isn’t. Mathematically, the circle is merely the bounded collapse of the spiral. It is complete, but life goes on’5.
Developing new ways to think about coral (left) and jellyfish (right, digitally inverted for clarity) life cycles, with use of layering, spiral shapes or continuous line drawings.
A final take-away from the workshop was that the process of drawing and expressing ideas with paper crafts also allowed participants to incorporate their own identities and personalities into their work. We generated a space for participants to step away from the regular routine of lab work, experiments and computational data analysis, and take the time to think more deeply about their research questions. Participants were encouraged to leave phones and social media behind, although we allowed their use to access relevant images and videos online, promoting a focused atmosphere throughout the workshop. Slowing down, reflecting and sharing imaginative time with colleagues through drawing, creating and discussion, has strong potential to lead to new insights into scientific questions. Workshops such as this could be one tool to help researchers actively engage with the often microscopic life they are studying, enabling a process-oriented approach to ‘flow, attend and flex’, as recently proposed by James Wakefield6. We recommend this style of workshop to other scientists searching for artistic ways of thinking about and communicating their work.
Acknowledgements
Thanks to the University of Exeter Living Systems Institute, for allowing us to host the workshop in their boardroom space, which was an ideal light-filled environment. This workshop was funded as part of a BBSRC David Phillips Fellowship (BB/T00990X/1) to Elizabeth Williams. We also thank each workshop participant for their valued contribution to this collaborative drawing workshop, which was in itself an experiment. Additional thanks to Dr Luis Bezares Calderónfor helpful comments on the text.
References
Anderson, G. 2014. Endangered: A study of morphological drawing in zoological taxonomy. Leonardo 47(3): 232 – 240.
Anderson-Tempini, G., 2017. Drawing as a Way of Knowing in Art and Science. Intellect Books.
Merian, M.S., Brafman, D. and Schrader, S., 2008. Insects & flowers: the art of Maria Sibylla Merian. Getty Publications.
Hodin, J., Ferner, M.C., Heyland, A., Gaylord, B., Carrier, T.J. and Reitzel, A.M., 2018. I feel that! Fluid dynamics and sensory aspects of larval settlement across scales. Evolutionary ecology of marine invertebrate larvae, 13, pp.190-207.
Gilbert, S.R. and Gilbert, S.F., 2023. “Process Epistemologies for the Careful Interplay of Art and Biology: An Afterword”, in Anderson-Tempini, G. and Dupré, J., 2023. Drawing Processes of Life: Molecules, Cells, Organisms, pg. 295.
Wakefield, James G. 2023. “Flow, Attend, Flex: Introducing a Process-Oriented Approach to Live Cell Biological Research”, in Anderson-Tempini, G. and Dupré, J., 2023. Drawing Processes of Life: Molecules, Cells, Organisms, pg. 280.
Open microscopy projects are flourishing with researchers contributing new technology and accessible workflows with the hope of democratising access to microscopy. In our upcoming webinar on Thursday 16 May at 15:00 BST, we’ll be hearing from Richard Bowman, Jan Huisken and Dumisile Lumkwana about three very different projects, OpenFlexure, Flamingo and VP-CLEM-KIT.
Richard will speak about OpenFlexure Microscopes, which are open-source optical microscopes that are built using 3D printed components and off the shelf components. You can read about the OpenFlexure microscope here.
Jan will discuss the Flamingo project. Flamingos are modular light sheet microscopy setups, which flip the concept of core microscopy facilities allowing the team to move custom advanced microscopes to the samples instead of taking samples to a core facility.
Dumi will discuss VP-CLEM-KIT, a new low-cost pipeline to support users to access high resolution volume correlative light-electron microscopy.
The deadline for entries for the FocalPlane-elmi204 has been extended until 21 May 2024. We would be delighted to receive entries from all imaging modalities, and you don’t need to be attending elmi2024 to enter. The winning entry, which will be selected by public vote following shortlisting by the elmi2024 organising committee, will be featured as the cover image on an upcoming issue of Journal of Cell Science.
When I was 25 and having a particularly bad day, crying to my mum about how exhausted and in pain I was during my studies, she recalled a passing comment made about my 10-year-old self by a homeopath who was treating my brother. The homeopath looked at me inquisitively, and then asked me if I could walk up and down the corridor at a comfortable pace. His response: “She’s just running on nothing, absolutely no energy reserve”. This man identified an issue 10 years prior to any diagnosis, and a further 10 years to being properly assessed.
And I am still an incredibly slow walker.
I have spent the past year, and indeed, my first-year postdoc salary, going through the private medical system in order to be diagnosed for a myriad of conditions, some of which I’ve had my whole life. This includes, but undoubtedly is not limited to: repetitive strain injury in both wrists, chronic migraines, fibromyalgia, hypermobility syndrome/EDS, degenerative lumbar spine, hip sclerosis, autism and anxiety. I am now a clinical trial comorbidity nightmare, a badge I wear with pride, if only as gallows humour. Over this time, I’ve had the opportunity to reflect on how on Earth I have managed to get through the education system, as well as work in multiple academic institutes. Now, in a far more supportive environment than I have ever worked in, I feel secure enough in my position to say my piece about why the current structure of wet lab science is nigh-on impossible for people with energy-limiting conditions or mobility issues, never mind the two.
I am not one to be afraid of ruffling feathers, and as such can say that having worked or studied at now six universities and institutes, some of which are the best in the UK, not one has shown significant change to improve the state of wet lab science’s innate ableism despite proclaiming “inclusivity”. I have worked on stem cells in one capacity or another for almost 10 years now, and I have yet to find one lab that is disability-accessible even for minor difficulties, even in recently built buildings. Lab benches have a top shelf that is so high up that I need to climb onto a chair, then the bench and then lean up to simply retrieve a DNA extraction kit. I’m 5’6 and one of the taller women in our lab; I am almost certain the standard lab benches were designed by the same individuals who forgot women wear seatbelts. The benches are non-adjustable and are often too narrow to fit a chair in, never mind a wheelchair. Ergonomic pipettes are regularly not available, and heavy lifting and copious bending is expected, just to simply collect what you need. I’ve lost count the number of times I have scooted on my bum on the floor to organise a drawer or to find a reagent in a freezer, like a dog that needs to go to the vet. A large amount of arm strength is required to open -80 freezers and lift cell storage shelves out of liquid nitrogen. Last time I checked, powerlifting was not an essential requirement on the job description. And don’t even get me started about requiring a cane in the lab. Despite the infrastructure remaining steadfastly unchanged, many colleagues have provided kind words and helpful hands over the years, but when discussing the sheer commitment of lab work, well, everybody seems to think we are all in the same boat.
Spoiler: we’re not.
Now I did not get into academia thinking it was rosy. Academic careers are famous for the dedication required, so much so that it puts off even the most privileged and enthusiastic of individuals. Similarly, stem cells are renowned for being needy: weekends, holidays, anniversaries, Christmas. This would wear down anyone, but for those with energy-limiting conditions, this spirals into a whole different beast leading to professionally difficult conversations. I’ve tried my best to be open about my energy limitations to my superiors in the past and been met with disdain masquerading as advice, being told that I was lying or that I should keep quiet as it makes me sound “weak”. I’ve had my conditions leveraged as a means of bribing for loyalty, and even had my pleas for consideration misinterpreted as vying for sympathy to get deadline extensions or funding, rather than just asking for basic human compassion. I’ve collapsed in the lab on multiple occasions, slumped on the floor from sheer exhaustion trying to meet unobtainable demands, secretly thinking that if I end up in hospital at least I’ll have “a valid excuse” to rest. The irony was I was in hospital at one point, and was still asked if I could attend a scheduled meeting. Although this very much was an extreme case of a toxic work environment, to a lesser extent I’ve observed similar behaviours in most institutes, either to myself or others, where the fear of backlash about disclosing or indeed managing one’s health issues is valid and can come with very real consequences.
All I can do is keep what advocation available to me small-scale, within my limits, just so I can still do my job.
So how do we tackle this issue, and indeed, support both current and new generations of scientists? I don’t know and nor do I feel it is the total responsibility of myself and the people who are affected by this multi-faceted, systematic monolith of an issue to resolve it, because we’re already tired. There are those who are sounding the horn plenty and still are ignored despite putting their all into it (and good on them!). For me, I’ve gained a surprising amount from being vocally open about all the weird and wonderful aches, pains and idiosyncrasies that I experience to colleagues and seniors, being unabashedly afraid to talk about it. What began as me just being honest about my life at one institute has snowballed into me being the local disability sign-poster at each institute I frequent. I have had so many wonderful interactions with people with similar or indeed completely different difficulties, asking for advice on deadlines, how to handle management, how to get accommodations, and frankly the most important, just having a sympathetic ear. On reflection, these interactions have paved a road for me to not feel immense guilt about my limited engagement and advocacy on a grander stage. Plus, I have a crumbling spine, pounding migraines, diminished energy and a shot nervous system, and have eventually admitted that I need to cut myself some slack.
What people don’t realise is that this is not an inspirational underdog story, because I live in constant fear there could be no happy ending. My academic career could end just as it’s begun, because I don’t know if my body can continue working in the lab for another 10 years until I’m office bound. I go home after a 9-5 day in the lab and can spend 2 hours in a bath just to be able to sleep that night. I take entire medical kits with me to conferences to make it through and still collapse at the end. I have got stuck at my desk because my legs have stopped working after being on my feet for 9 hours. If I don’t have lab work, I have to work at home, because open office spaces are not conducive to productivity when you are stimuli-sensitive. Like many postdocs, I am expected to maintain the juggling act of being both a full-time lab researcher and a trainee PI. The problem lies when you wake up already at 40% rather than 100%; this no longer is a “difficult period in one’s career that everyone goes through”, it’s the marathon from Hell and, as I mentioned, I’m a slow walker.
Even by writing this article and attaching my name to it, this could be found by committees and unconsciously (or indeed consciously) used to exclude me from funding opportunities.
My greatest fear is that those who called me weak were right, that I will not make it physically. However, I am not paving anything by being quiet, so this seems as accessible a vessel as I can manage. In short, if I were to provide some advice to others with a long list of chronic and acute conditions, it would simply be to be defiant. I won’t deny that I haven’t had an impeccable support network of friends and family, but even as a child, I have always vocally refused to believe that just because something is the way that it is does not mean it is right. Additionally, the best motivation you could give me is to say that I can’t do something. I often feel I should write some apology letters to my school teachers who were not expecting such defiance from an 8-year-old. This tenacity, however, is the only thing I feel got me to this point and is my advice to everyone, regardless of your situation. Demand your space, be seen, and be unafraid of how it will affect your career, because, for some such as myself, you may not even have one if your accommodations aren’t met.
Just keep reminding yourself that you are worth the effort and that you deserve to be in the room.
And For Managers, Principal Investigators and Senior Staff….
For PIs wishing to be more supportive and inclusive, I’d advise taking a leaf out of my current mentor’s book. This individual once said “I care more about the people than I do the cells” and shows it by their actions. They will come in on weekends to cover tissue culture, finish experiments if any of us call in sick, is happy to cover for me experimentally for any doctor’s appointments, if necessary, all while juggling their own childcare responsibilities. I am not afraid in the slightest to talk about how I really am, if I’m struggling, or if I require any additional help, although it took me almost a year to truly instil this trust. I had been upfront about my conditions prior to even applying for my position, and they have pushed for every accommodation I asked for from the get-go. Although I am aware of my sheer luck at finding such a considerate leader, this should not be the case. I honestly believe the only way attitudes will change is from the inside and by senior individuals supporting and advocating for those they are responsible for. After all, it takes a village to build a lab.
Although I’d be factually incorrect to say no progress has been made, what facet of academia do you think has grown the most in terms of disability inclusivity, and what leaves much to be desired?
The School is designed for Master’s students, PhD students and Postdocs who work in the field studying the Transcriptional Control of the Genome during Embryonic Development, Cell Differentiation, and Disease, or for those who would like to approach it.
One of the primary focuses will be to discuss how to combine “wet” and “dry” technologies to address computationally demanding, important biological questions.
A key feature of this course will be the low ratio between participants and teachers/speakers. This will allow each participant to come in close contact with successful scientists and learn how they developed key technologies to respond to pressing questions in their field.
The course will include talks, group discussions and group activities.
In addition to subject matter-specific topics introduced by professors, you will also benefit of hands-on analytical and computational sessions on:
Barcode-based Lineage Trancing
Understanding the 3D Genome
Transcription Factors and Peak Calling
in situ hybridization combined with scRNAseq
An important innovation of this year is that previous summer school participants, on the wave of their current scientific successes, have been selected to teach.
Participate and candidate yourself to being the next generation teacher!
The school will also include a short course on scientific writing for developmental and cancer biologists.
In this SciArt profile, we meet Philipp Dexheimer, who combines his research background and love for art to effectively communicate complex ideas in science to a broad audience. As an artist at heart and a scientist in mind, Philipp’s creations are inspired by the concepts and molecular aesthetics of nature. He works with diverse techniques and media, from Molecular Graffiti to scientific illustrations and videography.
C. elegans Graffiti in Vienna
Can you tell us about your background and what you work on now?
As a scientist, I have always been fascinated by the beauty and complexity of life. During my PhD at the IMP Vienna, I focused on RNA Biology, studying the functions of microRNAs in early animal development using C. elegans as a model system. For my Postdoc, I stayed with the nematodes but shifted my focus to research protein aggregation in the context of myopathies. Our results put protein misfolding on the map as a critical contributor to certain myopathies and suggest that caloric restriction is a promising treatment strategy in this context. My postdoc paper is about to be submitted, and I very recently left the lab to go all-in with Science Art, so it’s the beginning of an exciting new professional chapter for me.
Journal Covers for Molecular Cell, Genome Research and Embo Journal
Were you always going to be a scientist?
I always had a love for biology – Life is, after all, the most fascinating phenomenon in the universe – and the decision to study Molecular Biology after school came very intuitively without having to think about it much. Once I set foot in the lab for the first time as an undergrad, I figured that this is what I want to be doing. Next to the intellectual joy of being a detective investigating the intricacies of nature, the scientific community is just great company; one gets to meet so many inspiring people from all over the world. That’s what made me really feel at home in the institutes of this world, and I would not want to miss a single day of my time as an active researcher.
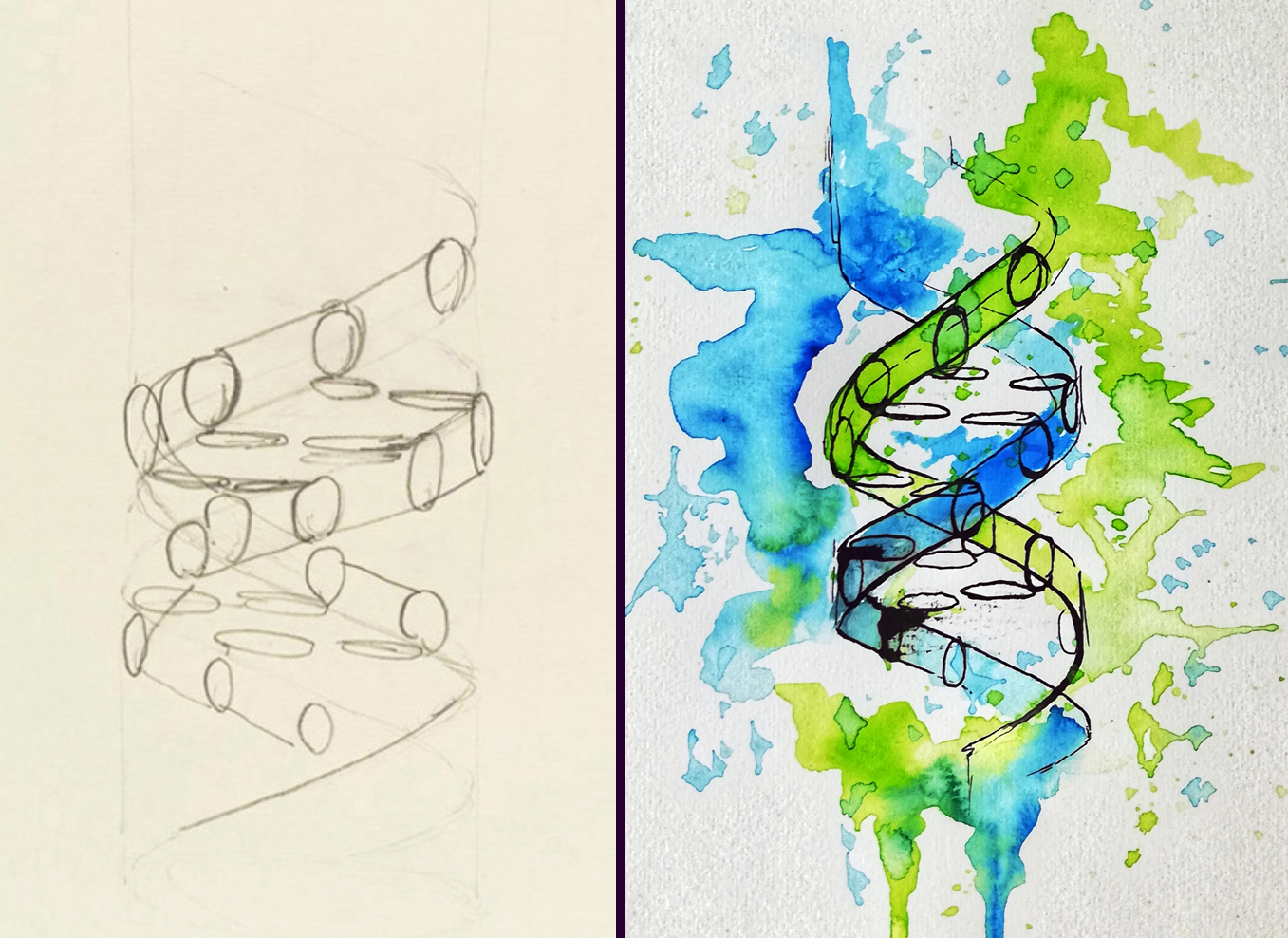
Re-imagining Francis Cricks’ DNA sketch using Ink and watercolor
And what about art – have you always enjoyed it?
In school, I always liked to doodle around when class wasn’t particularly interesting, but things didn’t start moving until I ended up picking up a spray can at the age of 15. I spent a lot of time at the skatepark and on basketball courts which, together with a general interest in Hip Hop culture, led me to start my artistic development by painting Graffiti. Since then, urban art continues to be a passion of mine. Once I got into the lab, I started to draw my inspiration more and more from biology, and by now the main topic of my creations revolves around nature.
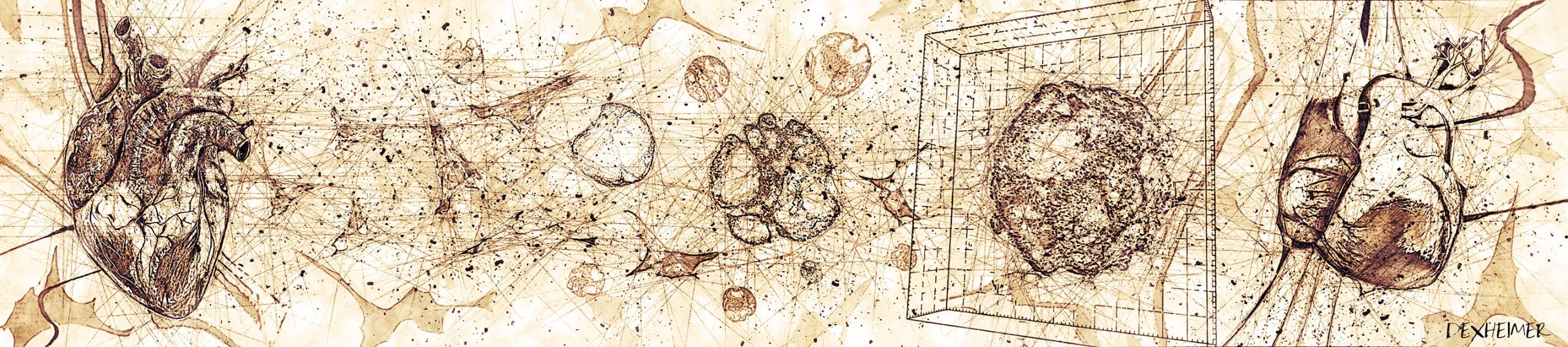
Heart organoids drawn in Leonardo Da Vinci’s style
What or who are your most important artistic influences?
My style has definitely been influenced a lot by urban art. In addition, I love comics and animated cartoons; they continue to be a source of inspiration for my work. More recently, I started to get into watercolor and ink drawings, which I love because the medium is very organic – just like biology itself. David Goodsell is one of my science art heroes; his paintings of the molecular world in scale are to me the most outstanding drawings of the molecular world.
Ubiquitin Graffiti in Vienna
How do you make your art?
When I don’t hold a spray can in my hand, I love working with Photoshop to create digital artwork for journal covers, scientific posters, or biotech homepages – finding a way to illustrate complex concepts in a way that resonates with the human mind & soul never fails to excite me. More recently, I also started to experiment with video creation and editing. Short videos are a great medium to make science accessible to a large audience, and I am convinced that they will become a more important part of the science communication toolbox in the future.
In addition, I think that AI tools are in the process of revolutionizing the way we design illustrations and other art forms – my digital workflows have become increasingly complex and by now involve hopping back and forth between many different programs. I would never take the direct output of any AI tool and use it as a final artwork, though. I want to have a personal touch in there, otherwise it just doesn’t feel like my own creation. AI-generated designs are mostly somewhat generic and lack the certain edge that makes art so interesting after all. My belief is that the interplay of AI prototyping and “classic” manual editing is the future of digital artwork.
Infographic depicting biodegradable spectral conversion films
Does your art influence your science at all, or are they separate worlds?
Both science and art draw a lot from the ability to extract the abstract essence of reality that manifests itself in concrete forms in the world surrounding it. While there is no direct influence of my artistic endeavors on the science I do in the lab, I think that training the cognitive ability of abstraction comes in handy when contemplating biology. In the end, great science aims not at describing a particular phenomenon in a certain organism which happens to be the subject of our studies, but rather at finding the bigger concepts behind the workings of Life.
What are you thinking of working on next?
There are many projects in the pipeline at the moment; my personal favorite goal for the upcoming years is to travel the world and paint large-scale murals on the walls of research institutes and public buildings. I think Molecular Graffiti represents a unique way of porting scientific concepts into public space and has a lot of potential to spark curiosity about the life sciences among a large audience, rendering biology accessible also to non-experts. So if you, dear Reader, happen to have an idea for a wall that could use a beautiful science mural, feel free to reach out!
What better place to hold a conference than a castle? Well, the 25th Conference of the German Society for Developmental Biology (GfE) was held in the historic castle of Osnabrück (see picture below) together with the Dutch Society for Developmental Biology (DSDB) and provided an excellent location to celebrate research. The four-day event brought together researchers from Germany, the Netherlands and around the world to share the latest findings and foster collaboration in the field of developmental biology, including areas such as stem cell biology, pattern formation, regeneration and disease.
The conference was held in the castle of Osnabrück.
Opened by the current president of the GfE, Prof. Kerstin Bartscherer, the meeting started with a keynote talk from Melina Schuh about the beginning of life ‘New insights into meiosis in mammalian oocytes’. After an unscheduled change to the program, due to various strikes and associated delays, and the pre-scheduled talk by Erez Raz on ‘The role of the Dead end protein in controlling the spatial organization and function of RNA molecules within zebrafish germ-cell granules’ the first two sessions ‘Stem cells and fate decisions’ gave broad overview about findings in stem cell dynamics in the plant Arabidopsis and in early mouse embryos, the specification during cardiac development and natural variation in cardiac regenerative capacity, nervous system development in Nematostella, and the skull development in the mouse. The sessions on stem cells were concluded by Jochen Wittbrodt, who gave a talk entitled ‘Towards the genetics of individuality’ using a population genetics approach to show genetic basis of quantitative cardiac phenotypes of two Japanese rice fish models. One of them has elevated heart rates associated with ventricular hypoplasia and impaired cardiac function, which may be related to loss-of-function mutations in candidate genes. For me, this was a highlight of the meeting, as it showed me once again that good research takes time.
The second day began with a special keynote lecture. 2024 is a very special year for developmental biologists: We celebrate the 100th anniversary of the Spemann-Mangold experiment. Christof Niehrs paid tribute to the famous Spemann-Mangold organization experiment and emphasized the importance of this discovery for developmental biology. Accordingly, the following sessions also dealt with ‘Emergence and maintenance of patterns’. Here, we heard how plants use MAP kinase signaling for early cell polarization and the generation of cell wall patterning in the plant vasculature, the self-organization of mucociliary epithelia in Xenopus and, in four insect talks, about feedback loops in the segmentation clock of the red flour beetle, as well as branching during neuronal dendrite differentiation, a role of the ECM receptor Dystroglycan is important for the blood-testis barrier formation and my own talk about basement membrane remodeling in organ formation, all using Drosophila.
In the afternoon we continued with the session ‘When location matters’ where we looked at the role of a RhoGEF in neural crest migration, mechanisms regulating cardiomyocyte invasion of collagenous tissue during zebrafish heart regeneration, apical constriction and cell polarity in cranial neural tube closure, and a proteomics approach to identify cell polarity regulators in plants. Later, the session ‘Genetic and epigenetic control of development’ illustrated how expression patterns are precisely regulated in time, new insights into the role of Pitx2 in cardiac pacemaker development and arrhythmogenesis, a novel function of the hox gene Antennapedia during muscle development, a neuronal subtype specification of spinal projection and motoneurons by a common temporal sequence, and the epigenetic regulation of seed development and plant speciation.
The third day began with a keynote by Susana Coelho on the origin, evolution, and regulation of sexual development through an “Algal views on evo-devo of sex determination”. The following section, “Quantifying and Modeling Development,” ranges from quantifications at the subcellular level, such as the specification of founder cells in lateral root formation, Semaphorin/Plexin Signaling in Collective Cell Migration or the orientation of microtubules in dendritic pruning, to single cell-resolution with studies on lineage-specific genetic modules during cranial development, muscle stem cell heterogeneity and alterations in the thymic niche to the development of reproductive tracts and even the regulation of whole body size. The section ‘Evolutionary adaptions’ took us to a journey across the genome size and gene family expansion in the genus Hydra and the β-catenin-driven endo-mesoderm specification as a Bilateria-specific novelty, but also how leaves adapt during evolution their phenotypes and the tolerance of mouse embryos on ectopic retroviral activity.
Of course, there is one thing that should not be missing at a conference. Day two and three were supplemented with extensive poster sessions in which scientific projects were hotly debated.
In a special session, three distinct awards were granted. Michael Brand holds the laudatory speech for Christiane Nüsslein-Volhard, who was honoured by the GfE with the Klaus Sander Lifetime Achievement Award. In an exploration of color and pattern in the animal kingdom, she elaborated on “Animal Beauty: Function and Evolution of Biological Aesthetics. She addressed the origin and relevance of what humans find beautiful in the animal kingdom. She introduced that color patterns in the animal kingdom have important functions in communication, e.g. in mate choice, but can also develop rapidly and with high variability, which is of great importance in terms of evolution, natural and sexual selection. We know relatively well how invertebrates develop their color patterns, but much is still unknown about vertebrates. Fish are interesting models for studying the development and evolution of color patterns in animals, because they have beautiful patterns made up of a mosaic of differently colored cells in the skin. Fittingly, since this work came from the Nüsslein-Volhard lab, the second award, the PhD award of the GfE goes to Marco Podobnik. In his talk “On the Genetic Basis of Pigment Pattern Diversification in Danio Fish” addressed the question which genes contribute to patterning differences between species. The third award, the GfE Hilde-Mangold Prize 2024, an award for young scientists, went to Daniel Wehner for his work on neuroregeneration after spinal cord injury. He showed how Small leucine-rich proteoglycans inhibit CNS regeneration by altering the structural and mechanical properties of the tissue in the lesion environment. In the evening, the networking event took place at a local nightclub, providing a top destination to make new connections. The poster award winners were also announced here.
The final day was opened by the keynote talk of Anna Akhmanova. She explained how microtubule dynamics control cell polarity and migration. The meeting was completed by two sessions addressing ‘Regeneration and disease models’ with talk that use organoids to model human heart or liver development, study cardiac injuries or regeneration in marine annelids, fish heart and the fish fin. Finally, Hugo Snippert talked about genetic heterogeneity in tumors and how he is studying this in patient-derived colon cancer organoids on the single-cell level.
The joint GfE/DSDB 2024 meeting was an absolute highlight for me. It always reminds me that the development of organisms is what interests me most in biology, and I strongly believe that developmental biology is the foundation for understanding human disease. I was lucky enough to attend the GfE meeting as a student and to have given a talk now as Postdoc is still incredible to me. I’m already looking forward to the 2026 GfE meeting in Potsdam.
Eloise Dries, Yannick Meyers, Daniel Liesner, Floriele Gonzaga, Jakob Becker, Eliane E Zakka, Tom Beeckman, Susana M Coelho, Olivier De Clerck, Kenny A Bogaert
Maria Oorloff, Adam Hruby, Maxim Averbukh, Athena Alcala, Naibedya Dutta, Toni Castro Torres, Darius Moaddeli, Matthew Vega, Juri Kim, Andrew Bong, Aeowynn J. Coakley, Daniel Hicks, Jing Wang, Tiffany Wang, Sally Hoang, Kevin M. Tharp, Gilberto Garcia, Ryo Higuchi-Sanabria
Devon E. Mason, Paula Camacho, Megan E. Goeckel, Brendan R. Tobin, Sebastián L. Vega, Pei-Hsun Wu, Dymonn Johnson, Su-Jin Heo, Denis Wirtz, Jason A. Burdick, Levi Wood, Brian Y. Chow, Amber N. Stratman, Joel D. Boerckel
Samantha M. Barnada, Aida Giner de Gracia, Cruz Morenilla-Palao, María Teresa López-Cascales, Chiara Scopa, Francis J. Waltrich Jr., Harald M.M. Mikkers, Maria Elena Cicardi, Jonathan Karlin, Davide Trotti, Kevin A. Peterson, Samantha A. Brugmann, Gijs W. E. Santen, Steven B. McMahon, Eloísa Herrera, Marco Trizzino
Dmitry A. Kretov, Leighton Folkes, Alexandra Mora-Martin, Noreen Syedah, Isha A. Walawalkar, Kim Vanyustel, Simon Moxon, George J. Murphy, Daniel Cifuentes
Isabel Zhang, Giulia LM Boezio, Jake Cornwall-Scoones, Thomas Frith, Ming Jiang, Michael Howell, Robin Lovell-Badge, Andreas Sagner, James Briscoe, M Joaquina Delás
Rossella Debernardis, Katarzyna Palińska-Żarska, Sylwia Judycka, Abhipsa Panda, Sylwia Jarmołowicz, Jan P. Jastrzębski, Tainá Rocha de Almeida, Maciej Błażejewski, Piotr Hliwa, Sławomir Krejszeff, Daniel Żarski
Keren Cheng, Yasunari Seita, Eoin C. Whelan, Ryo Yokomizo, Young Sun Hwang, Antonia Rotolo, Ian D. Krantz, Maninder Kaur, Jill P. Ginsberg, Priti Lal, Xunda Luo, Phillip M. Pierorazio, Rebecca L. Linn, Sandra Ryeom, Kotaro Sasaki
Stefano Comazzetto, Daniel L. Cassidy, Andrew W. DeVilbiss, Elise C. Jeffery, Bethany R. Ottesen, Amanda R. Reyes, Sarah Muh, Thomas P. Mathews, Brandon Chen, Zhiyu Zhao, Sean J. Morrison
Christopher Zdyrski, Vojtech Gabriel, Oscar Ospina, Hannah Wickham, Dipak K. Sahoo, Kimberly Dao, Leeann S. Aguilar Meza, Abigail Ralston, Leila Bedos, William Bastian, Sydney Honold, Pablo Piñeyro, Eugene F. Douglass, Jonathan P. Mochel, Karin Allenspach
Olivia Sniezek Carney, Kodi William Harris, Yvonne Wohlfarter, Kyuna Lee, Grant Butschek, Arianna Anzmann, Steven M Claypool, Anne Hamacher-Brady, Markus Andreas Keller, Hilary J Vernon
V. Pragathi Masamsetti, Nazmus Salehin, Hani Jieun Kim, Nicole Santucci, Megan Weatherstone, Hilary Knowles, Jane Sun, Riley McMahon, Josh B Studdert, Nader Aryamanesh, Ran Wang, Naihe Jing, Pengyi Yang, Pierre Osteil, Patrick P.L Tam
Xin-Min Li, Hannah Jenke, Sören Strauss, Yi Wang, Neha Bhatia, Daniel Kierzkowski, Rena Lymbouridou, Peter Huijser, Richard S. Smith, Adam Runions, Miltos Tsiantis
Ranjita Thapa, Karl H. Kunze, Julie Hansen, Christopher Pierce, Virginia Moore, Ian Ray, Liam Wickes-Do, Nicolas Morales, Felipe Sabadin, Nicholas Santantonio, Michael A Gore, Kelly Robbins
Florian Laurent, Simon Maria Bartsch, Anuj Shukla, Felix Edgardo Rico Resendiz, Daniel Couto, Christelle Fuchs, Joel Nicolet, Sylvain Loubery, Henning J Jessen, Dorothea Fiedler, Michael Hothorn
Bastienne Zaremba, Amir Fallahshahroudi, Céline Schneider, Julia Schmidt, Ioannis Sarropoulos, Evgeny Leushkin, Bianka Berki, Enya Van Poucke, Per Jensen, Rodrigo Senovilla-Ganzo, Francisca Hervas-Sotomayor, Nils Trost, Francesco Lamanna, Mari Sepp, Fernando García-Moreno, Henrik Kaessmann
Shatha Salameh, Devon Guerrelli, Jacob A. Miller, Manan Desai, Nicolae Moise, Can Yerebakan, Alisa Bruce, Pranava Sinha, Yves d’Udekem, Seth H. Weinberg, Nikki Gillum Posnack
Semih Bayraktar, James Cranley, Kazumasa Kanemaru, Vincent R Knight-Schrijver, Maria Colzani, Hongorzul Davaapil, Jonathan Chuo Min Lee, Krzysztof Polanski, Laura Richardson, Claudia Semprich, Rakeshlal Kapuge, Monika Dabrowska, Ilaria Mulas, Shani Perera, Mina Patel, Yen Ho, Xiaoling He, Richard Tyser, Laure Gambardella, Sarah Teichmann, Sanjay Sinha
James Cranley, Kazumasa Kanemaru, Semih Bayraktar, Vincent Knight-Schrijver, Jan Patrick Pett, Krzysztof Polanski, Monika Dabrowska, Ilaria Mulas, Laura Richardson, Claudia Semprich, Rakeshlal Kapuge, Shani Perera, Xiaoling He, Siew Yen Ho, Nadav Yayon, Liz Tuck, Kenny Roberts, Jack Palmer, Hongorzul Davaapil, Laure Gambardella, Minal Patel, Richard Tyser, Sanjay Sinha, Sarah Teichmann
This is part of the ‘Lab meeting’ series featuring developmental and stem cell biology labs around the world.
Where is the lab?
The Welshhans lab is located at the University of South Carolina, which is in Columbia, South Carolina, USA.
Research summary
The Welshhans Lab works on neural development. In particular, we are interested in the process by which neural connectivity is formed. This process is mediated by a highly dynamic sensory and motor structure located at the ends of developing axons, called the growth cone. Much of our work focuses on local translation, which is the process by which a subset of mRNAs is localized to and locally translated within growth cones to regulate axon guidance. We study how this molecular mechanism and others regulate typical development. Furthermore, we study how the dysregulation of various molecular mechanisms, including local translation and adhesion, contribute to the phenotypes of Down syndrome. We use mouse models and human-induced pluripotent stem cell-derived neurons and brain organoids to study these processes.
Can you give us a lab roll call?
Katelyn, PhD Candidate: My project examines the local translation of b-actin and how it regulates axon guidance through adhesion-based mechanisms during nervous system development.
Nikita Kirkise, PhD candidate: I am a 4th year PhD student in Kristy Welshhans’ lab. My project investigates the role of extracellular matrix proteins, specifically laminins, in regulating the local translation of mRNAs in growth cones of developing mouse cortical neurons.
Jordan Headen, PhD candidate: I am a second-year PhD student investigating the role of adhesion and the local translation of candidate mRNAs during the development of the nervous system.
We also have three undergraduates in the lab who are studying how adhesion and local translation are altered in Down syndrome. For their research, they are using human fibroblasts as a model.
Favourite technique, and why?
Kristy: My favorite technique is anything involving microscopy and living cells. I find it fascinating to watch living cells under high magnification. It doesn’t matter whether we are using brightfield, fluorescent translation reporters, or some other fluorescent tagging method, I can stare at these movies for hours and always find something interesting!
Apart from your own research, what are you most excited about in developmental and stem cell biology?
Kristy: I am most excited about some recent advances that are improving the quality of life for individuals with neurodevelopmental disorders (including non-pharmacological, pharmacological, gene therapy, and stem cell-based treatments). Many disorders still have no treatment, but recent advances in some of these areas are opening new doors that I hope will continue to gain momentum.
How do you approach managing your group and all the different tasks required in your job?
Kristy: I am a pretty organized person and have a never-ending (but prioritized) to-do list, so that helps me stay on top of things. Every Friday, I time block my calendar with all the activities I need to accomplish in the week ahead. In addition, I meet one-on-one with everyone in the lab every week. Overall, the most important thing to me is the success of my lab members, which means something different for each individual. So, I prioritize that and then fit in everything else around it!
What is the best thing about where you work?
Kristy: In our Department, there is a very supportive group of four faculty who all work on axon biology. This makes it an optimal environment not only for me but for my lab as well. I am also part of a larger group at the University, which is the Carolina Autism and Neurodevelopment (CAN) Research Center. This is a multidisciplinary group composed of faculty and their labs that study neurodevelopment and related disorders. We are composed of people from very diverse disciplines, including Biology, Psychology, Public Health, Computer Science, etc., which has allowed me to think and collaborate with others on my research in novel ways.
Katelyn: The supportive and collaborative environment of both the lab itself, and throughout the University.
Nikita: The best thing about my work is the lab itself. We have such supportive and fun lab members (including our super supportive mentor) that it makes the PhD journey a little less daunting.
Jordan: I like the collaborative and supportive nature of the Welshhans Lab and the department as a whole.
What’s there to do outside of the lab?
Kristy: I love that Columbia is only two hours from the beach and two hours from the mountains. I love to go mountain biking, hiking, and generally spend time outdoors with my family all year round.
Katelyn: Columbia has many parks and lakes to enjoy and is not far from beautiful beaches and the Blue Ridge Mountains. Columbia also has great breweries to explore while enjoying the nice weather.
Nikita: Columbia has many parks and trails, most of which are along the riverside, where you can catch the most beautiful sunsets. We also have a lot of restaurants to try around the University.
Jordan: The Riverwalk is nice for walking and biking throughout the whole year and tubing during the summer. Folly Beach is also enjoyable around the summer months for being in the water and trying the different restaurants near the beach.
In May, we celebrate the winners of Development’s 2023 Outstanding Paper Prize, hearing from the authors of two papers describing the roles of the Frizzled receptor. Chaired by Development’s Executive Editor, Katherine Brown.
Tuesday 21 May – 15:00 BST
Ling Loh (The George Washington University) Joe Hanly (The George Washington University and Duke University) ‘From Signaling to Patterning: Dissecting the roles of Frizzled receptors in butterfly wing development’
Megan Michalski (Van Andel Institute) ‘Successful therapeutic intervention in new mouse models of frizzled 2-associated congenital malformations’
At the speakers’ discretion, the webinar will be recorded for viewing on demand. To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
 (No Ratings Yet)
(No Ratings Yet)



 (1 votes)
(1 votes)