Our lab is located on the 4th floor of the biomedical building at University of California, Santa Cruz. We are also affiliated with the Institute for the Biology of Stem Cells and Genomics Institute.
Research summary
Ali Shariati: We are interested in understanding unifying principles of stem cell fate decisions during the early days of mammalian development. We also use CRISPR-based approaches to control stem cell fate decisions and assemble them into embryo-like structures.
Lab role call
Gerrald Lodewijk, Postdoctoral Fellow
Research focus: Human embryonic development and implantation, transcriptional mechanisms of G0 to G1 transition in mammalian cell division cycle.
Ben Topacio, Postdoctoral Fellow
Research focus: Determining molecular mechanism linking between cell cycle and cell fate decision in pluripotent stem cells.
Abolfazl Zargaro, PhD student
Research Focus: Development of tools for quantitative analysis of single-cell live microscopy data.
Silvart Arabian, PhD Student
Research Focus: How cells decide to re-enter the cell cycle when they are in G0 phase. Transcriptional mechanisms of G0 to G1 transition in mammalian cell division cycle.
Seungho Lee, PhD Student
Research Focus: How MAPK signalling shape the chromatin landscape and its dynamics in pluripotent stem cells.
Undergraduate researchers: Sayaka Kozuki (Synthetic Models of the embryos), Clara Han (G0 to G1 transition), Lucy Suarez (Cell Cycle Control in pluripotent cells), Arshia Kapil (Microscopy image analysis), Shivani Rao (Microscopy image analysis), Dami Ibrahim (Microscopy Image Analysis).
Favourite technique, and why?
Ali: Biological systems are quite dynamic and time is implicit in biological transitions such as cell fate decisions. So, my favorite method is Single Cell Live Microscopy because it allows us to capture dynamics of cell fate decisions with high temporal resolution. Live microscopy allows us to directly watch individual stem cells as they make important decisions such as to differentiate or not to differentiate. With live microscopy, one has access to information such as morphology of cells (shape, size…), spatial relations of cells and dynamics of signalling pathways that is typically lost in other single cell approaches. Occasionally, you also find cases of individual cells that behave completely different than what is described in the literature.
I am also interested in using CRISPR-based devices to control cell fate and cellular response to signalling cues.
Apart from your own research, what are you most excited about in developmental and stem cell biology
Ali: I am pretty excited about collective cellular decision making that results in reproducible formation of patterns during development as well as application of synthetic biology approaches to control collective behaviour of cells.
How do you approach managing your group and all the different tasks required in your job?
Ali: Managing an academic research laboratory has different aspects such as how to manage projects or how to build a scientifically vibrant environment. One thing I have done is to take an adaptive approach because scientific endeavours are inherently very dynamic in nature and require flexibility in planning. As a result, I am constantly revising my plans and I guess I am also still learning how to lead an academic research laboratory. Periodically, I reflect on how to improve my mentorship and how to promote a dynamic scientific culture that allows for training future generations of scientists with genuine and deep interest in discovering new biology. That last statement is actually one of my professional dreams!
What is the best thing about where you work?
Ali: The campus is so beautiful with many redwood trees, ocean views and wildlife. My colleagues are very supportive and understanding and I also have the chance to work with some very motivated students.
Seungho: Supportive environment, not cutthroat; the opportunity to do research on unanswered questions on stem cells; the Bay area is the centre of innovation in biotech, providing more opportunities to collaborate with other labs or companies.
Silvart: A chance to surround yourself with experts from various research fields. Gain exposure to new technologies and broaden your understanding of stem cells and cell cycle regulation. A supportive environment that fosters growth and development.
Gerrald: Commuting to the lab by bike and getting a free workout! Building our postdoc community (UCSC Postdoc Association).
Ben: I like the lab environment. Everyone is helpful and supportive.
Sayaka: A great opportunity to have research experience during the undergraduate
What’s there to do outside of the lab?
Ali: What I do outside the lab varies a lot. I enjoy (very) long walks with my family, friends or sometimes on my own. I set learning goals for myself such as learning a new language. I also enjoy helping students from resource-limited regions of the Middle East to find their way into higher education.
Seungho: Nature activities; Golf; Santa Cruz doesn’t offer much in terms of nightlife or city activities (great for those who prefer a more peaceful, quiet environment), but San Jose/Palo Alto are not too far away.
Silvart: Sports, playing music and reading books. Spending time outside in nature.
Gerrald: Go to the climbing gym and sauna afterwards. Take a walk or bike ride in one of the nearby state parks.
Ben: Go home. The SF bay area is a short drive away.
Sayaka: Visiting Monterey Aquarium, going Asian supermarkets.
Browse through other ‘Lab meeting’ posts featuring developmental and stem cell biology labs around the world.
A new edition of the annual Young Embryologist Network conference (YEN 2023) will be hosted at the Francis Crick Institute, in London, on the 22nd of May 2023.
YEN conferences, organised by PhD students and postdocs, have massively increased their impact since virtual attendance was offered in 2021. We are committed to providing an opportunity for early-career developmental biologists to share their research, network, and engage with colleagues in the field. Thus, a hybrid format will also be held this year and attendance will be free thanks to the support of our sponsors.
YEN 2023 will cover some of the most exciting topics in the field, including cross-species comparisons, germ-cell development, and stem-cell-based models of development among others. We have the pleasure of welcoming Andrea Pauli (IMP, Vienna), Madeline Lancaster (MRC LMB, Cambridge), Katsuhiko Hayashi (Osaka University), and Henrik Kaessmann (ZMBH, Heidelberg) as invited speakers. Prof. Henrik Kaessmann will give the Sammy Lee memorial keynote address.
Moreover, the highly acclaimed “Scientific Perspectives” talks are back. This year, the session will focus on “Equality, Diversity and Inclusion in Science”. Rafael Galupa (CBI, Toulouse) and Alison Forbes (Head of the Equality, Diversity & Inclusion committee at the Crick, London) will share their perspectives and experiences on this important subject.
PhD students and postdocs who want to give a short talk or present a poster may submit an abstract by the 20th of April 2023. Registration will be open until the 9th of May 2023.
You can register for this conference, and read a little more about our community, in the link below:
This year’s BSDB Annual General Meeting (AGM) will be held during the Sheffield Spring focus meeting on Cellular Plasticity in Morphogenesis. It will take place on Tuesday the 18th of April, as 16:15 in the main lecture theatre of the conference venue (The Edge). If you would like to attend virtually, please use this link: BSDB AGM Zoom Meeting
Meeting ID: 979 7241 4300
Passcode: dWx@Y7
We would also like to remind you that we will be electing three new BSDB committee members. Please send nominations together with the names of the proposer and seconder to the BSDB Secretary (secretary@bsdb.org) by the 14th of April 2023. Instructions on how to vote will be provided during the AGM.
We look forward to seeing you all there!
Finally, registration opens very soon for the BSDB-hosted European Developmental Biology Congress 25th-28th September 2023. Keep an eye on our website for more info.
If you look at, especially say, identical twins, if one person does testing, you are automatically finding out for your twin what their genetic status is. And so that’s tough, right? Because you’re deciding for them.
Kira Dineen, host of DNA Today
In the latest episode of the Genetics Unzipped podcast, we’re discussing Chris Hemsworth’s recent Alzheimer’s risk findings and the pros and cons of direct-to-consumer DNA testing with genetic counsellor and host of the podcast DNA Today, Kira Dineen.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
We recently announced that we will be working with three newly appointed the Node correspondents, who will be helping us to develop and write content for the Node in 2023. We caught up with each of them to chat about their research backgrounds and the topics that they’re excited to write about over the course of the coming year.
First up is Brent Foster, a technician at the Whitney Laboratory for Marine Bioscience, University of Florida, where he is studying the evolutionary origin of nervous systems using a range of marine invertebrates. As you will see, Brent has a longstanding interest in creative writing alongside his scientific career and began to blend the two by writing for his local newspaper. We talked about how his experience of using unconventional experimental organisms influences his writing, and the importance of being part of a science writing community.
Congratulations on being selected as a the Node correspondent. Why did you decide to apply for the role?
I decided to apply for it mostly because I had such a good experience with The Company of Biologists’ Creative Science Writing Workshop that I went to last year. I respect the mission of The Company of Biologists and their whole philosophy, and I thought this would be a really cool opportunity to expand beyond the readership I’ve had so far with my science writing.
What led you to first become interested in science writing? And what sort of science writing have you been doing so far?
I’ve always liked science. My dad was a high school biology teacher, and we lived in a rural area. I got to know the local wildlife, local trees and all of that fairly well. I really enjoyed the outdoors and I’ve always loved learning how nature works. And then on the flip side, once I started my undergraduate degree, I discovered that I enjoyed writing as well. And I had never really thought of pairing the two – I just considered writing as a hobby and assumed science would be my career, and that was my understanding going forward.
When I started working in the lab at the UF Whitney Laboratory, I met up with a local news editor. We struck up a conversation and I found out he was interested in science, but didn’t have a science background, and I was interested in writing, but hadn’t really done any formal writing, other than a few personal essays in college. And he basically issued a challenge to write something for the Palm Coast Observer, which is the local newspaper here. So, I approached my supervisor, and I asked him if it would be ok to write up some articles about what the lab is doing for the newspaper. From there, I discovered that I really enjoyed writing about the sciences, maybe even a little bit more than I enjoyed doing the experiments in the laboratory, and so I’ve expanded from there. The past two years I’ve presented some work at the Society for Integrative and Comparative Biology (SICB) meeting, and there I met some people who write for the SICB blog, and so I’ve contributed a few posts there. Last year I applied to the Creative Science Writing Workshop, hosted by The Company Biologists, loved it, and I’ve kind of snowballed into larger and larger projects. In October, I went to a science writing conference in Memphis, where I met a few folks and learned a little bit about the life of a science writer.
Thinking about what you were saying there about going to workshops and conferences focused on science writing, have you found that meeting likeminded scientists who also have an interest in creative writing has influenced you?
Yeah, absolutely. That was one of the big things from the Workshop last year: I was kind of shocked at how all of these brilliant scientists really just seemed to want to have some sort of creative outlet. And not always just about the work that they’re doing themselves, which showed me how creative science and scientists can be. So that was really eye opening. And then in January, when I was at SICB, I announced that I was a correspondent for the Node. I had a couple of people come up to me afterwards and we were throwing ideas back and forth about some different writing projects. That was a lot of fun. So, it has opened up some doors as well as friendships that I don’t think I would have found otherwise.
In terms of your own research, what has your career path been so far? And what’s your current research focus?
This seems to be the story for a lot of people now, but my career path is very, very windy. When I got started as an undergraduate student, I wanted to be an epidemiologist. After my first semester, I deferred my education and lived in Brazil for two years. I learned the language and the richness of that culture, and after just two months I noticed that I started dreaming in Portuguese. When I returned to my University, I attended a club meeting for deaf and hearing-impaired students. I only know rudimentary American Sign Language (ASL), so I’m sure my fingers were just fumbling with an accent. To offset the clumsiness of my hands, I would mouth the words I was trying to spell. It was only after the meeting when I realised that the words I mouthed were in Portuguese, not English. Somehow ASL and Portuguese had gotten tangled up in my brain, and both of them seemed sectioned off from English. I thought that was pretty fascinating. So, in the end, my undergraduate degree was in neuroscience, with minors in linguistics and creative writing.
I was especially interested in neurolinguistics, so how language is processed in the brain. That was my attempt to try to marry my interest in writing and science. I did some eye tracking studies, MRI studies, and EEG. And as I was getting all these great research experiences, I was a little disappointed in how little we could actually tell from those particular methods. I mean, you can tell a lot by each of them, but there were no clear causal effects, right? It was all “well, this is happening, and we are interpreting it to mean maybe this.” It wasn’t very definite, and that was a little disappointing for me when trying to understand some basic biology.
Then my wife was accepted to a PhD programme at the University of Florida, and so we moved down here. Originally, we were on the main campus, and I was in an MRI lab where we did a lot of cognitive neuroscience research related to attention and anxiety. They were really, really cool projects, and I’m grateful for the experience that I had there, but again, I had the same feeling of, “man, I’m just not sure how much we can actually tell from what we’re doing.” My wife and I stayed on main campus for the first year of her PhD. And then she moved out here to the Whitney Lab, and I didn’t want to have to commute back and forth. So, I actually reached out to the director of the lab, Mark Martindale (who’s now my supervisor), and I told him about my research experience. I had no wet lab experience – all of the studies that I’d done were human studies and nothing at all relevant to what they do here at the Whitney Laboratory. But I was interested in learning and asked if I could come down even just on the weekends to get some wet lab experience. And he turned around and offered me a job, and that’s how I started here.
So, I’ve switched from neurolinguistics to basic marine biology and developmental biology. And I love it. I feel like I can understand the outcomes of the experiments that I do a little bit clearer than I could within the broader cognitive sciences. I don’t mean to disparage other sciences as I think there are a lot of valuable things that we can learn from them – it just wasn’t for me.
Do you find that the science you’re doing now is more inspiring for your writing than what you were studying before? Because I imagine the sort of organisms you’re working on are a lot more interesting from that creative science writing perspective.
Yeah, that’s a great question. I think people could relate more to what I was doing before, when I was asking questions like “what’s the brain doing?” and “how do people process language?” In the eye-tracking lab, I came to understand a little bit about how people read, which has been helpful in the technical part of crafting my writing in a way that’s easy to read. But thinking back to what I found cool about science when I was a kid, I was always drawn to weird, quirky animals that do weird, quirky things – nature’s a real creative playground. Now I get to tell people that I work with sea anemones, and they’ll say, “I don’t know if I know anyone who works with a sea anemone,” or I mention that I work with comb jellies, and they say, “oh, what’s a comb jelly?” That’s a really fun type of interaction, to hear people saying, “that’s so cool. I never knew that.” And I try to capture that fun as much as I can in my writing.
Thinking about the sort of things that you’re going to be writing for the Node, what topics are you excited to write about? And do they relate to your own work or are you going to be branching out a bit into other areas?
Well, like I said, the quirky biology is what gets me excited, so I’m hoping to focus on non-model organisms, organisms that not everyone knows or hears about. This will probably involve a little bit of the work in our lab, because there are a lot of us here at the Whitney Laboratory who work with non-model organisms that are great for answering very specific questions but often get glossed over just because they’re not so well established.
My secondary focus is related to the idea of working with non-model organisms. Because these organisms are not as well established, I want to highlight labs that have to develop their own technologies or develop their own techniques or adapt existing techniques for their animals. This is hard and I think a lot of times underappreciated, both by the public and even by other scientists. With Drosophila, for example, you can do all sorts of things which you might take for granted. By contrast, I just talked to a scientist a few weeks ago who is trying to adapt transgenic approaches for cuttlefish, and they’ve spent almost a year trying to get something to work. So, I want to show those two aspects of something that I don’t think gets shown a lot, or at least doesn’t get into the major headlines. I want to give non-model organisms their due, I guess!
What are you hoping to gain from the experience of being a the Node correspondent?
Most of my science writing so far has been either hyperlocal or very specific to one group of biologists from a single conference, so one thing I’m hoping to gain as a correspondent for the Node is reaching a broader audience. I know the Node is pretty popular internationally, and I haven’t written for an international audience before. So I hope to stretch myself a bit in that area. I am looking forward to meeting other scientists in my field, hearing their stories and helping to share those stories.
I am also excited to interact with the Node team and other correspondents to learn about the craft of science writing a bit more. I don’t really have any formal training in science communication so I’m looking forward to participating in workshops. And of course, I’ve mentioned this several times now, but just having a community is huge. I didn’t realise how big the science writing community is until last year, and it’s gratifying to feel welcomed to it. Before I discovered it, I almost felt like an outsider. I was always asking myself, “is there a place for these two different aspects of my identity that everyone else seems to think of as a contradiction?” It’s fun to see that there are indeed other people who are combining their interests in science and writing.
Apart from writing, what do you enjoy doing in your spare time?
Lately, I have been doing a lot of writing! But I do enjoy the outdoors, particularly kayaking or canoeing – there are some lovely springs around here. In fact, I have a kayak trip this weekend and I’m hoping to see some manatees. I enjoy playing the piano, too. Primarily classical piano, though I don’t get a lot of opportunities to do that these days. And I enjoy reading. As a student, you kind of feel as if you either don’t have the time to read or that sometimes you’re reading about things that you’re not too excited about. In the gaps between studies, it’s been fun to be able to read some things just for the pleasure of reading.
Do you find that this sampling of other people’s writing has an influence on your own work?
Yeah, absolutely. Because I’ve gotten so interested in science writing, a lot of what I’ve been reading lately is by other science writers. That’s been informative in learning how other people approach science writing, but I also enjoy regular literature. I’ll pick up a piece of fiction or even poetry sometimes. And to be honest the poetry reading that I do, and the creative writing classes I took as an undergraduate that taught me a little bit about poetry, have actually informed my ability to write concisely in a way that I’m not sure I would have picked up from other genres. So, I think that by dabbling a little bit in a novel here, a short story there, some poetry, all of those things, you can take something from each one of those genres that together can make a science writing piece pretty powerful.
At the Gurdon Institute, which is part of the University of Cambridge. We are located in the city centre of Cambridge, UK.
Research summary:
EmmaRawlins: We ask, how are our lungs built and maintained? We have a particular focus on stem cell biology of the developing and adult lungs and asking how cell-cell interactions regulate growth, patterning and repair. We have also recently started collaborating with human geneticists to explore the genetic contribution to chronic lung diseases in humans, and whether these have their origins in infancy, or the adult, or both.
Rawlins lab, Christmas party 2022
Lab role call:
Claire: 1st year PhD student investigating airway homeostasis in humans.
Emma: PI – does all the paperwork.
Hannah: Clinical lecturer – has completed an MD and a PhD and is now balancing postdoctoral work with clinical duties, working on lineage decisions in the developing airway.
John: Postdoc interested in morphogenesis of the developing human lung.
Kyungtae: Postdoc interested in cell-cell signalling during lung alveolar development.
Odara: 1st year PhD student jointly supervised by Vito Menella who is interested in analysis of volumetric electron microscopy and using human organoids to study toxicology.
Quitz: Lab manager who keeps us all organised and enjoys thinking about computational biology methods.
Tessa: PhD student working on epithelial cell fate decisions in the developing mouse lung, and optimal imaging techniques for visualizing lung development.
Vanesa: PhD student (just submitted her thesis!) jointly supervised by Kristian Franze and works on the role of stiffness in cell fate decision making during human lung development.
Yihong: Visiting final year PhD student from Zhejiang University who is interested in tuft cells.
Ziqi: PhD student working on the role of hypoxia in cell fate decision making during normal human lung development.
Favourite technique, and why?
Emma: Clonal lineage-tracing combined with genetic manipulation and microscopy, and now often coupled to single cell transcriptomics. In my opinion, this is the most elegant method to study normal and aberrant cell fate decisions due to the presence of mutant and control cells in the same tissue. One current lab challenge is how to apply this technique to human samples.
Apart from your own research, what are you most excited about in developmental and stem cell biology
Emma: The advances being made in understanding of regeneration from organisms that regenerate on an impressive scale like Axolotls.
How do you approach managing your group and all the different tasks required in your job?
Emma: Most of the time it feels like neither managing my group, or the other admin and teaching tasks, are being done properly. It’s important to recognise that good-enough is sufficient for many tasks we must do, particularly the administrative ones. I try and prioritise my lab as it’s the science that excites me. I block time out in my diary to ensure that I have a detailed science conversation with every lab member every 2 weeks, and other tasks are fitted in around this schedule. I also have annual career planning meetings with everyone in the lab (the University’s appraisal scheme) and try and make sure that I keep up to date with any changing aspirations in addition. That way I/we can be looking out for the best career development opportunities for each person.
What is the best thing about where you work?
Claire: Working in Cambridge offers so much rich scientific history alongside cutting-edge techniques. It’s incredible to train in a such a collaborative space, and the Rawlins lab has been such a supportive and kind group with lots of advice and guidance as I begin my PhD.
Emma: Cambridge has a really rich scientific environment, it’s very easy to network to find collaborators in pretty much any research area that you are interested in.
Kyungtae: The best place in the world to do science – especially developmental biology. Also, it’s easy to get connected between fields to fields and person to person, leading to fantastic interdisciplinary studies.
Tessa: The lab environment is very supportive and collaborative, it’s an exciting place to do science and work with like-minded colleagues who are happy to help you troubleshoot or discuss new ideas.
What’s there to do outside of the lab?
Claire: As a PhD student, I’m part of a college here at Cambridge, and you have the opportunity to meet peers across different subject areas over weekly dinners and different social events. My favourite thing to do is explore Cambridge by going on runs around town and trying all the different restaurants!
Kyungtae: There are great walks and places around the Cambridge with beautiful nature which I can feel the universe – that is why it is the University of Cambridge, in Cambridge. Work and walk and feel the Cambridge.
Tessa: Cambridge has a lot of green space, and (weather permitting!) you can spend lots of time outdoors on the meadows or walking through the backs of the Colleges and admiring the architecture.
Browse through other ‘Lab meeting’ posts featuring developmental and stem cell biology labs around the world.
We are delighted to announce a new series on the Node called ‘Lab meetings’! In these posts, we will be highlighting developmental and stem cell biology labs across the globe. We aim to build up a directory of labs, which will not only showcase the exciting research and researchers in the community, but will also provide a useful resource for anyone looking for their next job. We ask the group leaders about their research and mentoring style, while the group members share what they like about the lab and what they like to do away from work.
Our first ‘Lab meeting’ is with the Rawlins lab, at the Gurdon Institute in Cambridge, which you can read here. You’ll be able to see all our full directory on our ‘Lab meetings‘ page.
Drop us an email if you would like to nominate a lab (including your own), especially if you have (or will soon have) open positions. We look forward to working with you all to build up a useful resource!
It’s not ridiculous to suggest that the Y chromosome might eventually become so mutation-addled that it disappears entirely. In fact, it’s already happened…in the Amami spiny rat.
Please join us for the International Symposium on Women in Tunicata Biology. The Tunicata are invertebrate chordates, several of which are model organisms in developmental biology (e.g. Ciona intestinalis, Botryllus schlosseri). The symposium will be Tuesday, March 28 and Wednesday, March 29, starting at 6 am Pacific Time/9 am Eastern Time. Speakers will be honoring the work of retired researchers and presenting their own research. All researchers are welcome to attend. Please email Marie Nydam (mnydam@soka.edu) or Anna Di Gregorio (adg13@nyu.edu) for a schedule and Zoom link.

 (No Ratings Yet)
(No Ratings Yet)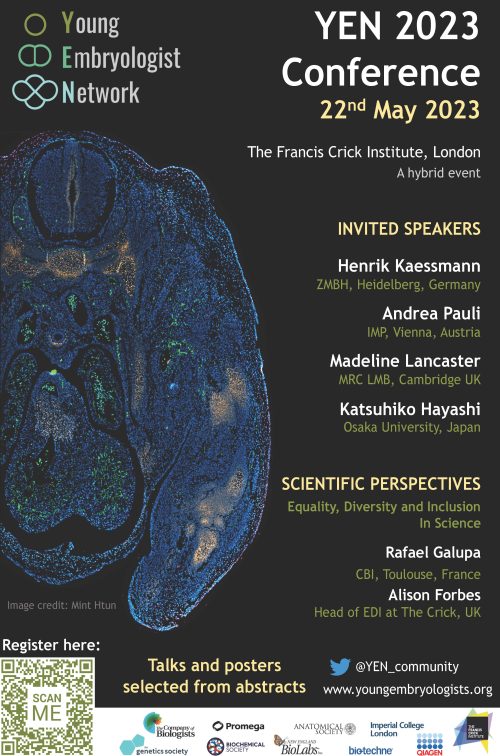
 (1 votes)
(1 votes)



